Das könnte Ihnen auch gefallen
- Acta Horticulturae 17 Growth of Sugarloaf' Pineapple On Mine Tailings Amended With Mineral Soil and Palm Mesocarp FiberDokument8 SeitenActa Horticulturae 17 Growth of Sugarloaf' Pineapple On Mine Tailings Amended With Mineral Soil and Palm Mesocarp FiberJeffry Ortiz GamboaNoch keine Bewertungen
- Control of Flower Formation and Fruit Size in The PineappleDokument11 SeitenControl of Flower Formation and Fruit Size in The PineappleJeffry Ortiz GamboaNoch keine Bewertungen
- A Genetic Map of Pineapple (Ananas Comosus (L.)Dokument16 SeitenA Genetic Map of Pineapple (Ananas Comosus (L.)Jeffry Ortiz GamboaNoch keine Bewertungen
- Cell Wall Alterations in The Leaves of Fusariosis-Resistant and Susceptible Pineapple Cultivars PDFDokument9 SeitenCell Wall Alterations in The Leaves of Fusariosis-Resistant and Susceptible Pineapple Cultivars PDFJeffry Ortiz GamboaNoch keine Bewertungen
- Acta Horticulturae 17 Growth of Sugarloaf' Pineapple On Mine Tailings Amended With Mineral Soil and Palm Mesocarp FiberDokument8 SeitenActa Horticulturae 17 Growth of Sugarloaf' Pineapple On Mine Tailings Amended With Mineral Soil and Palm Mesocarp FiberJeffry Ortiz GamboaNoch keine Bewertungen
- Characterization of A Field-Grown Transgenic Pineapple Clone Containing The Genes Chitinase, AP24, and BarDokument7 SeitenCharacterization of A Field-Grown Transgenic Pineapple Clone Containing The Genes Chitinase, AP24, and BarJeffry Ortiz GamboaNoch keine Bewertungen
- Acta Horticulturae 12 Pre - and Postharvest Metabolism of Crown Leaves of Pineapple FruitDokument6 SeitenActa Horticulturae 12 Pre - and Postharvest Metabolism of Crown Leaves of Pineapple FruitJeffry Ortiz GamboaNoch keine Bewertungen
- Acta Horticulturae 15 Field Evaluation of Mechanization System For Large Scale Pineapple Production On Mineral Soils in MalasyaDokument8 SeitenActa Horticulturae 15 Field Evaluation of Mechanization System For Large Scale Pineapple Production On Mineral Soils in MalasyaJeffry Ortiz GamboaNoch keine Bewertungen
- Acta Horticulturae 16 Effect of Planting Density On Growth, Development and Yield of Irrigated Pineapple in Nghe An ProvinceDokument6 SeitenActa Horticulturae 16 Effect of Planting Density On Growth, Development and Yield of Irrigated Pineapple in Nghe An ProvinceJeffry Ortiz GamboaNoch keine Bewertungen
- Acta Horticulturae 13 Leaf Gas Exchange of Pineapple As Influenced by FruitDokument6 SeitenActa Horticulturae 13 Leaf Gas Exchange of Pineapple As Influenced by FruitJeffry Ortiz GamboaNoch keine Bewertungen
- Acta Horticulturae 14 The Effect of NaCl On The Mineral Nutrient and Photosynthesis Pigments Content in Pineapple in Vitro PlantetsDokument8 SeitenActa Horticulturae 14 The Effect of NaCl On The Mineral Nutrient and Photosynthesis Pigments Content in Pineapple in Vitro PlantetsJeffry Ortiz GamboaNoch keine Bewertungen
- Acta Horticulturae 01 Rejuvenating The Malaysian Pineapple IndustryDokument14 SeitenActa Horticulturae 01 Rejuvenating The Malaysian Pineapple IndustryJeffry Ortiz GamboaNoch keine Bewertungen
- Acta Horticulturae 16 Effect of Planting Density On Growth, Development and Yield of Irrigated Pineapple in Nghe An ProvinceDokument6 SeitenActa Horticulturae 16 Effect of Planting Density On Growth, Development and Yield of Irrigated Pineapple in Nghe An ProvinceJeffry Ortiz GamboaNoch keine Bewertungen
- Acta Horticulturae 12 Pre - and Postharvest Metabolism of Crown Leaves of Pineapple FruitDokument6 SeitenActa Horticulturae 12 Pre - and Postharvest Metabolism of Crown Leaves of Pineapple FruitJeffry Ortiz GamboaNoch keine Bewertungen
- Acta Horticulturae 04 Pineapple Breeding For Quality Improvement in South VietnamDokument8 SeitenActa Horticulturae 04 Pineapple Breeding For Quality Improvement in South VietnamJeffry Ortiz GamboaNoch keine Bewertungen
- Acta Horticulturae 10 F1 Hybrid Pineapple Resistant To Bialaphos HerbicideDokument8 SeitenActa Horticulturae 10 F1 Hybrid Pineapple Resistant To Bialaphos HerbicideJeffry Ortiz GamboaNoch keine Bewertungen
- Acta Horticulturae 11 Sunn Hemp Cover Cropping and Solarization As Alternatives To Soil Fumigants For Pineapple ProductionDokument12 SeitenActa Horticulturae 11 Sunn Hemp Cover Cropping and Solarization As Alternatives To Soil Fumigants For Pineapple ProductionJeffry Ortiz GamboaNoch keine Bewertungen
- Acta Horticulturae 06 Sugar Accumulation Difference Between The Various Sections During Pineapple DevelopmentDokument10 SeitenActa Horticulturae 06 Sugar Accumulation Difference Between The Various Sections During Pineapple DevelopmentJeffry Ortiz GamboaNoch keine Bewertungen
- Acta Horticulturae 07 Changes in Sucrose Content and Related Enzyme Activities During Pineapple Inflorecence DevelopmentDokument8 SeitenActa Horticulturae 07 Changes in Sucrose Content and Related Enzyme Activities During Pineapple Inflorecence DevelopmentJeffry Ortiz GamboaNoch keine Bewertungen
- Acta Horticulturae 08 Evaluation of A Bioassay For Screening Resistance of Pineapple To RootDokument8 SeitenActa Horticulturae 08 Evaluation of A Bioassay For Screening Resistance of Pineapple To RootJeffry Ortiz GamboaNoch keine Bewertungen
- Acta Horticulturae 03 Pineapple Breeding Fulfilling Expectations of The Global Supply ChainDokument6 SeitenActa Horticulturae 03 Pineapple Breeding Fulfilling Expectations of The Global Supply ChainJeffry Ortiz GamboaNoch keine Bewertungen
- Acta Horticulturae 02 Pineapple Production and Research in ChinaDokument8 SeitenActa Horticulturae 02 Pineapple Production and Research in ChinaJeffry Ortiz GamboaNoch keine Bewertungen
- Acta Horticulturae 05 Identification and Characterization of Differentially ExpressedDokument10 SeitenActa Horticulturae 05 Identification and Characterization of Differentially ExpressedJeffry Ortiz GamboaNoch keine Bewertungen
- Acta Horticulturae 09 Sugar Accumulation in Smooth Cayenne' Pineapple Fruits in Different Harvest SeasonsDokument8 SeitenActa Horticulturae 09 Sugar Accumulation in Smooth Cayenne' Pineapple Fruits in Different Harvest SeasonsJeffry Ortiz GamboaNoch keine Bewertungen
- Temperature Determines The Occurrence of CAM or C3 Photosynthesis in Pineapple Plantlets Grown in VitroDokument7 SeitenTemperature Determines The Occurrence of CAM or C3 Photosynthesis in Pineapple Plantlets Grown in VitroJeffry Ortiz GamboaNoch keine Bewertungen
- Effect of Night Temperature On Flowering and Fruit Size in PineappleDokument4 SeitenEffect of Night Temperature On Flowering and Fruit Size in PineappleJeffry Ortiz GamboaNoch keine Bewertungen
- Canons of Pro Ethics-Standards of Conduct Licensees 2017Dokument3 SeitenCanons of Pro Ethics-Standards of Conduct Licensees 2017Jeffry Ortiz GamboaNoch keine Bewertungen
- How To Write A Business ... R Investors - ChronDokument3 SeitenHow To Write A Business ... R Investors - ChronJeffry Ortiz GamboaNoch keine Bewertungen
- Crias de La Tortuga Laúd Dermochelys CorDokument13 SeitenCrias de La Tortuga Laúd Dermochelys CorJeffry Ortiz GamboaNoch keine Bewertungen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (120)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
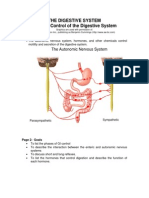
- ControlDokument12 SeitenControlSirVietaNoch keine Bewertungen
- Aetiology, Pathology and Management of Enterocutaneous FistulaDokument34 SeitenAetiology, Pathology and Management of Enterocutaneous Fistularoselinekhadija100% (1)
- Bioenergetics 1Dokument30 SeitenBioenergetics 1suni alditaNoch keine Bewertungen
- NCP 1Dokument1 SeiteNCP 1hsiriaNoch keine Bewertungen
- Pseudo ExfoliationDokument6 SeitenPseudo ExfoliationvannyanoyNoch keine Bewertungen
- Boerhaave Syndrome: A Case Report: Maj William B. Marshall, CRNA, MS, MAE, USAF, NC Beavercreek, OhioDokument4 SeitenBoerhaave Syndrome: A Case Report: Maj William B. Marshall, CRNA, MS, MAE, USAF, NC Beavercreek, OhioВукосав Н. ТркуљаNoch keine Bewertungen
- 5CRMSDokument2 Seiten5CRMSpandoraelitesNoch keine Bewertungen
- Kriya For Pelvic Balance PDFDokument2 SeitenKriya For Pelvic Balance PDFDiana Sánchez GómezNoch keine Bewertungen
- Bio Quiz BeeDokument2 SeitenBio Quiz BeeRuchi Briam James LagitnayNoch keine Bewertungen
- Bp503t Pcol Unit-VDokument46 SeitenBp503t Pcol Unit-VAakkkNoch keine Bewertungen
- PhysioEx Exercise 9 Activity 5Dokument3 SeitenPhysioEx Exercise 9 Activity 5Gonzeilo11Noch keine Bewertungen
- Pacemaker Invasive Cardiac PacingDokument57 SeitenPacemaker Invasive Cardiac PacingAhmad Khalil Ahmad Al-SadiNoch keine Bewertungen
- Theories of Impression Making in Complete Denture TreatmentDokument19 SeitenTheories of Impression Making in Complete Denture Treatmentdrgayen6042100% (1)
- The Effect of Nacl 0.9% and Nacl 0.45% On Sodium, Chloride, and Acid - Base Balance in A Picu PopulationDokument7 SeitenThe Effect of Nacl 0.9% and Nacl 0.45% On Sodium, Chloride, and Acid - Base Balance in A Picu Populationzack dahlanNoch keine Bewertungen
- Mumbai AyurvedaDokument2 SeitenMumbai AyurvedaAditya SinghNoch keine Bewertungen
- Client Screening Website FormDokument2 SeitenClient Screening Website FormBrett Whipp100% (2)
- Principles of Endocrinology Szathmari Miklos 2010Dokument25 SeitenPrinciples of Endocrinology Szathmari Miklos 2010Ryan James Lorenzo MiguelNoch keine Bewertungen
- Histochemistry and Enzyme HistochemistryDokument39 SeitenHistochemistry and Enzyme Histochemistrykay100% (2)
- Desolation - RL CaulderDokument308 SeitenDesolation - RL CaulderInes LagarrigueNoch keine Bewertungen
- PsychopharmacologyDokument77 SeitenPsychopharmacologychachaazkaNoch keine Bewertungen
- Haemoptysis Diagnosis and TreatmentDokument57 SeitenHaemoptysis Diagnosis and TreatmentMuhammad Cholid AlfahroziNoch keine Bewertungen
- Patterns of Electron Flow Through Light Reaction EventsDokument25 SeitenPatterns of Electron Flow Through Light Reaction EventsMar'J0% (1)
- Phonetics & Phonology (Danial Nadeem, Areeba Hamid, Shahrose Shahzadi, Muhammad Dawood)Dokument9 SeitenPhonetics & Phonology (Danial Nadeem, Areeba Hamid, Shahrose Shahzadi, Muhammad Dawood)Danial GujjarNoch keine Bewertungen
- Human Anatomy 4th Edition McKinley Test Bank 1Dokument35 SeitenHuman Anatomy 4th Edition McKinley Test Bank 1donnawugnwsjrzcxt100% (27)
- Class XI Practicals Term2Dokument5 SeitenClass XI Practicals Term2Prince KNoch keine Bewertungen
- InterneuronsDokument6 SeitenInterneuronsRafu DestajoNoch keine Bewertungen
- Aging Changes in The Heart and Blood VesselsDokument4 SeitenAging Changes in The Heart and Blood VesselsTotalenesya Reforrent SutiknoNoch keine Bewertungen
- Before Diving Make A Complete Dive Plan Together & Estimate Sea ConditionsDokument8 SeitenBefore Diving Make A Complete Dive Plan Together & Estimate Sea ConditionsMike LuckyNoch keine Bewertungen
- VEALDokument8 SeitenVEALcutiepattotieNoch keine Bewertungen
- START Triage Training PresentationDokument85 SeitenSTART Triage Training PresentationAnita Amanda100% (1)