Das könnte Ihnen auch gefallen
- RESUMEN Inmunidad AntibacterianaDokument6 SeitenRESUMEN Inmunidad AntibacterianaLetycia GodoyNoch keine Bewertungen
- 12 Resistencia A Bacterias y HongosDokument5 Seiten12 Resistencia A Bacterias y HongosValeria Dos SantosNoch keine Bewertungen
- Respuesta Inmune Frente A BacteriasDokument28 SeitenRespuesta Inmune Frente A BacteriasClaudia Echevarria SantosNoch keine Bewertungen
- InmunologíaDokument10 SeitenInmunologíaevangelineNoch keine Bewertungen
- Integridad de La PielDokument11 SeitenIntegridad de La Pielsonia eliana chozo mestanzaNoch keine Bewertungen
- Actividad3 MariaferzavalalaraDokument9 SeitenActividad3 MariaferzavalalaraMARIA FERNANDA ZAVALA LARANoch keine Bewertungen
- Mecanismo de Patogenicidad BacterianaDokument3 SeitenMecanismo de Patogenicidad BacterianaKeyla Vente VelascoNoch keine Bewertungen
- Inmunidad Frente A Bacterias ExtracelularesDokument10 SeitenInmunidad Frente A Bacterias ExtracelularesLaura RojasNoch keine Bewertungen
- Inmunidad Contra HongosDokument11 SeitenInmunidad Contra HongosMaryori Treyci ILasaca ValenzuelaNoch keine Bewertungen
- InmunologíaDokument12 SeitenInmunologíaMaria LópezNoch keine Bewertungen
- Inmunidad Celular Seminario 5Dokument8 SeitenInmunidad Celular Seminario 5Blanca CastilloNoch keine Bewertungen
- Inmunidad Frente A Bacterias, Virus y HongosDokument39 SeitenInmunidad Frente A Bacterias, Virus y HongosDavid Vasquez50% (2)
- E. ColiDokument5 SeitenE. ColiDaniela GonzálezNoch keine Bewertungen
- Resumen Fagocitos y Linfocitos NKDokument9 SeitenResumen Fagocitos y Linfocitos NKJoshein PerezNoch keine Bewertungen
- Inmunidad Frente A Bacterias Intracelulares - SalmonellaDokument9 SeitenInmunidad Frente A Bacterias Intracelulares - SalmonellaBrian Misari AnchiraicoNoch keine Bewertungen
- Inmunidad Frente A BacteriasDokument6 SeitenInmunidad Frente A BacteriasOscar GarzaNoch keine Bewertungen
- Apuntes de Anatomia Del S. Respiratorio y DigestivoDokument14 SeitenApuntes de Anatomia Del S. Respiratorio y DigestivoPau GallardoNoch keine Bewertungen
- Defensa Inmunitari1Dokument4 SeitenDefensa Inmunitari1Lesly Villca riosNoch keine Bewertungen
- 1-El Sistema Imunológico y La Inmunidad InnataDokument24 Seiten1-El Sistema Imunológico y La Inmunidad InnataYolandaNoch keine Bewertungen
- Micro II Respuesta Inmune A BacteriasDokument14 SeitenMicro II Respuesta Inmune A BacteriasDaniel MoraNoch keine Bewertungen
- Protozoosis Transmitidas Por Fecalismo. PatogenicidadDokument8 SeitenProtozoosis Transmitidas Por Fecalismo. PatogenicidadFranco RomeroNoch keine Bewertungen
- T-11 Inmunología Nuevo 2014Dokument37 SeitenT-11 Inmunología Nuevo 2014Sonia GarciaNoch keine Bewertungen
- Inmunidad Antibacteriana 2016-1Dokument39 SeitenInmunidad Antibacteriana 2016-1Kiki CarrilloNoch keine Bewertungen
- Respuesta InmuneDokument12 SeitenRespuesta InmuneVictoria DarritchonNoch keine Bewertungen
- 4°-Guía de Estudio - Sistema InmuneDokument16 Seiten4°-Guía de Estudio - Sistema InmuneDanilo Enrique Correa RosaNoch keine Bewertungen
- BROTE S Aureus Resistente A OxacilinaDokument4 SeitenBROTE S Aureus Resistente A OxacilinaMartin TarapuesNoch keine Bewertungen
- Reacciones HipersensibilidadDokument9 SeitenReacciones HipersensibilidadIvan Orellana De Leon100% (4)
- Fisio Tema 30Dokument8 SeitenFisio Tema 30lucisNoch keine Bewertungen
- Seminario 4 PDFDokument8 SeitenSeminario 4 PDFnahuel piscorzNoch keine Bewertungen
- Patogenia de Las Enfermedades PeriodontalesDokument7 SeitenPatogenia de Las Enfermedades PeriodontalesSandra Delgado100% (1)
- LupusDokument14 SeitenLupusKarla EscotoNoch keine Bewertungen
- Informe 07 - Inmunología Sem - Grupo 19Dokument15 SeitenInforme 07 - Inmunología Sem - Grupo 19Jenny TarrilloNoch keine Bewertungen
- Mecanismos de Algunas Bacterias para Evadir La Respuesta Inmune EXPO INMUNODokument3 SeitenMecanismos de Algunas Bacterias para Evadir La Respuesta Inmune EXPO INMUNOJimena ReyesNoch keine Bewertungen
- Tema 4 Respuesta Inmune Frente A Bacterias (Con Nuevo Punto)Dokument16 SeitenTema 4 Respuesta Inmune Frente A Bacterias (Con Nuevo Punto)luisdonielNoch keine Bewertungen
- Tema 4. Respuesta Inmunitaria Frente A BacteriasDokument9 SeitenTema 4. Respuesta Inmunitaria Frente A BacteriasKatheryne DesiderioNoch keine Bewertungen
- Inmunidad Adaptativa Frente A Las Bacterias ExtracelularesDokument7 SeitenInmunidad Adaptativa Frente A Las Bacterias ExtracelularesJoshue MurrietaNoch keine Bewertungen
- FagocitosisDokument16 SeitenFagocitosisJeisonAlejandroÁlvarezVergaraNoch keine Bewertungen
- PatogenicidadDokument7 SeitenPatogenicidadShirley vanessa Vega BedoyaNoch keine Bewertungen
- Atlas de InmunologiaDokument50 SeitenAtlas de InmunologiaNuñez Fonseca Jose100% (1)
- Tema 20Dokument7 SeitenTema 20sandraNoch keine Bewertungen
- Analizar Las Modalidades de Acción PatógenaDokument7 SeitenAnalizar Las Modalidades de Acción Patógenanayeli chilanNoch keine Bewertungen
- Cuestionario Orientación TP N°9 PatogeniaDokument4 SeitenCuestionario Orientación TP N°9 PatogeniaValentina BenavidezNoch keine Bewertungen
- Adaptaciones ParasitariasDokument4 SeitenAdaptaciones ParasitariascriznahpNoch keine Bewertungen
- Tipos de InmunidadDokument3 SeitenTipos de InmunidadMari GzlezNoch keine Bewertungen
- Inmunidad Frente A BacteriasDokument55 SeitenInmunidad Frente A BacteriasKarely IreneNoch keine Bewertungen
- Respuesta Inmune Frente A Bacterias IntracelularesDokument2 SeitenRespuesta Inmune Frente A Bacterias Intracelulares熊解Noch keine Bewertungen
- EstafilococcoDokument5 SeitenEstafilococcoVictoria DominioniNoch keine Bewertungen
- FAGOCITOSISDokument29 SeitenFAGOCITOSISMarcell Aquino ArboledaNoch keine Bewertungen
- Respuesta Inmunitaria Por Cristhian Tomsich Romo-LerouxDokument17 SeitenRespuesta Inmunitaria Por Cristhian Tomsich Romo-LerouxCristhianNoch keine Bewertungen
- Cap 01Dokument13 SeitenCap 01Fernando UsedaNoch keine Bewertungen
- Inmunidad Frente A PatógenosDokument6 SeitenInmunidad Frente A PatógenosamɩʀaɴdaNoch keine Bewertungen
- 10 Salmonella TyphiDokument8 Seiten10 Salmonella TyphiAlder Rios GoicocheaNoch keine Bewertungen
- InmunidadDokument12 SeitenInmunidadLulu PerezNoch keine Bewertungen
- Microbiología Humana N 12 GonorreaDokument50 SeitenMicrobiología Humana N 12 GonorreaLuis Gonzales RodriguezNoch keine Bewertungen
- Capitulo 17Dokument4 SeitenCapitulo 17Dalton CedeñoNoch keine Bewertungen
- Zambrano Mendoza Maite-PeriodonciaDokument11 SeitenZambrano Mendoza Maite-PeriodonciaMaite Lindsay ZambranoNoch keine Bewertungen
- Preguntas Finales Resueltas Del Sistema InmunologicoDokument13 SeitenPreguntas Finales Resueltas Del Sistema InmunologicoErick Silva100% (1)
- InmunoDokument8 SeitenInmunoAndres ErasoNoch keine Bewertungen
- Ssr-Rotafolio 1 PDFDokument41 SeitenSsr-Rotafolio 1 PDFmagdalena piña67% (3)
- Prueba de Biología 7º AñoDokument3 SeitenPrueba de Biología 7º AñoXimena Vásquez MelgarejoNoch keine Bewertungen
- Anatomía Topográfica Del CorazónDokument3 SeitenAnatomía Topográfica Del CorazónMariana Requena100% (2)
- Músculos Del Miembro Superior-Hombro y BrazoDokument4 SeitenMúsculos Del Miembro Superior-Hombro y BrazoSarai GomezNoch keine Bewertungen
- Ciclo Estral de La CabraDokument7 SeitenCiclo Estral de La Cabraleonel milanoNoch keine Bewertungen
- Somatico Vs AutonomoDokument55 SeitenSomatico Vs AutonomoValentina FernandezNoch keine Bewertungen
- CLASE 10 Y 11 Aparato Digestivo I (1) AnatomiaDokument56 SeitenCLASE 10 Y 11 Aparato Digestivo I (1) Anatomiavictor quispe CastroNoch keine Bewertungen
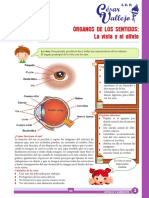
- Ojo Resumen MooreDokument9 SeitenOjo Resumen MooreLaura MinieriNoch keine Bewertungen
- Músculos de La RespiraciónDokument24 SeitenMúsculos de La RespiraciónNahuel CaballeroNoch keine Bewertungen
- Práctica de Epitelio de Revestimiento, Glándular y Conectivo. 2018Dokument19 SeitenPráctica de Epitelio de Revestimiento, Glándular y Conectivo. 2018Nicole BravoNoch keine Bewertungen
- Zona DT y VTDokument51 SeitenZona DT y VTGerald D. FrizNoch keine Bewertungen
- U.D. 2. Sistema Tegumentario. La Piel. Higiene y Aseo Del Paciente. Úlceras Por Presión.Dokument30 SeitenU.D. 2. Sistema Tegumentario. La Piel. Higiene y Aseo Del Paciente. Úlceras Por Presión.jusreadNoch keine Bewertungen
- Deglución en AdultoDokument11 SeitenDeglución en AdultoAndrea Gayoso GarridoNoch keine Bewertungen
- Cya3° - Iii BimDokument24 SeitenCya3° - Iii Bimeleyvac eclcNoch keine Bewertungen
- Pie Equinovaro CongenitoDokument19 SeitenPie Equinovaro CongenitoJair MachucaNoch keine Bewertungen
- Células Madre y Progenitoras para La Reparación de Cartílago ArticularDokument12 SeitenCélulas Madre y Progenitoras para La Reparación de Cartílago ArticularAMY ANDREA ARMIJOS DUTANNoch keine Bewertungen
- Latarjet SeminarioDokument43 SeitenLatarjet SeminarioDIEGO JOSUE MANRIQUE CASTILLONoch keine Bewertungen
- Rev. Salud Mov. 2015 7 (1) : 19-32: ISSN 2027-4548Dokument14 SeitenRev. Salud Mov. 2015 7 (1) : 19-32: ISSN 2027-4548Emily DavilaNoch keine Bewertungen
- Anemia Celulas FalciformesDokument9 SeitenAnemia Celulas FalciformesMayara OliveiraNoch keine Bewertungen
- Insuficiencia CardiacaDokument19 SeitenInsuficiencia CardiacaCarlos PalaciosNoch keine Bewertungen
- Libro Neurofisiologia de La Conducta Ebook 2015Dokument480 SeitenLibro Neurofisiologia de La Conducta Ebook 2015KarenArias88% (8)
- Borobia 5 027 033 02Dokument8 SeitenBorobia 5 027 033 02yoseph medranoNoch keine Bewertungen
- Tumores OMSDokument54 SeitenTumores OMSAna Maria CobosNoch keine Bewertungen
- Malformaciones Del Aparato Genital FemeninoDokument17 SeitenMalformaciones Del Aparato Genital FemeninoAndre Dario Ferandez100% (1)
- Cell Biology of Prokaryotic Organelles - En.esDokument19 SeitenCell Biology of Prokaryotic Organelles - En.esAlejandra arecasNoch keine Bewertungen
- Alteraciones de Los Cuerpos Formes de La SangreDokument35 SeitenAlteraciones de Los Cuerpos Formes de La SangredavideflNoch keine Bewertungen
- Hemoglobinas TalasemiasDokument95 SeitenHemoglobinas Talasemiasminichuper100% (1)
- Enfermedades HematologicasDokument63 SeitenEnfermedades HematologicasITZEL GUADALUPE GARCIA REYESNoch keine Bewertungen
- U1 CASO CLÍNICO DE MANEJO PREOPERATORIO EN CIRUGÍA PEDIÁTRICADokument8 SeitenU1 CASO CLÍNICO DE MANEJO PREOPERATORIO EN CIRUGÍA PEDIÁTRICAJhonatan Francis Rengifo ChotaNoch keine Bewertungen
- QUEILOSDokument22 SeitenQUEILOSpaul christian muñoz barrancoNoch keine Bewertungen