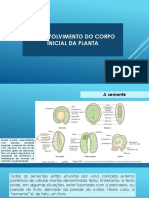
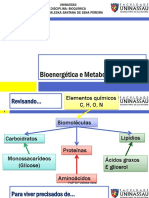
Das könnte Ihnen auch gefallen
- My Arquivo Germinação de Sementes PDFDokument110 SeitenMy Arquivo Germinação de Sementes PDFviniciusolssonNoch keine Bewertungen
- Relatório Fisiologia Da GerminaçãoDokument9 SeitenRelatório Fisiologia Da GerminaçãoDébora Lemes FacciNoch keine Bewertungen
- Sistema Vegetativo Apresentação Sobre SementesDokument14 SeitenSistema Vegetativo Apresentação Sobre SementesJAMILE O MONDEGONoch keine Bewertungen
- Temática - Ciências da Vida - MicrorganismoVon EverandTemática - Ciências da Vida - MicrorganismoNoch keine Bewertungen
- Multiplicação de Plantas OrnamentaisDokument37 SeitenMultiplicação de Plantas OrnamentaisAntónio Figueira100% (1)
- Fisiologia Da GerminacaoDokument21 SeitenFisiologia Da GerminacaoLúMacedoNoch keine Bewertungen
- BiologiaDokument12 SeitenBiologiaferjumomutombeneNoch keine Bewertungen
- 09043418082016morfologia e Anatomia Vegetal Aula 12Dokument10 Seiten09043418082016morfologia e Anatomia Vegetal Aula 12Doka Victor VictorNoch keine Bewertungen
- Texto de Apoio Nº2Dokument37 SeitenTexto de Apoio Nº2hghghg1Noch keine Bewertungen
- Germinação e Dormência SementesDokument43 SeitenGerminação e Dormência SementesCleber Jadoski100% (1)
- 1 Fisiologia VegetalDokument3 Seiten1 Fisiologia VegetalObsanjoNoch keine Bewertungen
- Ib Ufu MorfvegetalorgaSEMENTEDokument5 SeitenIb Ufu MorfvegetalorgaSEMENTELeandro Inácio LeiteNoch keine Bewertungen
- Semente - Wikipédia, A Enciclopédia LivreDokument10 SeitenSemente - Wikipédia, A Enciclopédia LivrenhashajaniNoch keine Bewertungen
- GerminaçãoDokument41 SeitenGerminaçãoLuziane TrovãoNoch keine Bewertungen
- SEMENTE MaterialDokument7 SeitenSEMENTE MaterialJone Alide QuissadoNoch keine Bewertungen
- f13827896 AVALIA O 2Dokument6 Seitenf13827896 AVALIA O 2laylaNoch keine Bewertungen
- Grad Unidade XI - Reprodução em Plantas SuperioresDokument14 SeitenGrad Unidade XI - Reprodução em Plantas Superiorescarliane_rebecaNoch keine Bewertungen
- Grad Unidade XIII - Dormência e GerminaçãoDokument19 SeitenGrad Unidade XIII - Dormência e Germinaçãocarliane_rebecaNoch keine Bewertungen
- Reproducao Nos Seres VivosDokument14 SeitenReproducao Nos Seres VivosRicardinhorickNoch keine Bewertungen
- Biologia - Reinos VegetaisDokument13 SeitenBiologia - Reinos VegetaisNatália ZardoNoch keine Bewertungen
- Cultura Do Milho - IntroduçãoDokument44 SeitenCultura Do Milho - IntroduçãoKeli Libardi RamellaNoch keine Bewertungen
- 5-Biologia VegetalDokument168 Seiten5-Biologia VegetalMayara StefanyNoch keine Bewertungen
- 3.5. Correlaciones Crecimiento - G-LDokument42 Seiten3.5. Correlaciones Crecimiento - G-LNATALIA ARIANA ALBUJAR RODRIGUEZNoch keine Bewertungen
- FT Reproduc3a7c3a3o AssexuadaDokument5 SeitenFT Reproduc3a7c3a3o Assexuadabiogeo1011Noch keine Bewertungen
- Germinação e Dormência de Sementes FlorestaisDokument22 SeitenGerminação e Dormência de Sementes FlorestaisRoberson VieiràNoch keine Bewertungen
- A SementeDokument9 SeitenA SementeJessuyn MatsinheNoch keine Bewertungen
- 2 Apostila ForragiculturaDokument8 Seiten2 Apostila Forragiculturathales de moura lopesNoch keine Bewertungen
- Material Propagação Sexual EnviarDokument11 SeitenMaterial Propagação Sexual EnviarCarlos André RamosNoch keine Bewertungen
- Biologia - Reino Vegetal Felipe FrancoDokument4 SeitenBiologia - Reino Vegetal Felipe FrancofesfeNoch keine Bewertungen
- Te T PDFDokument6 SeitenTe T PDFPedro Luis DomingosNoch keine Bewertungen
- Teste 12º PTAS - Reprodução Assexuada Nos Seres Vivos (1) - CópiaDokument6 SeitenTeste 12º PTAS - Reprodução Assexuada Nos Seres Vivos (1) - CópiaIsaura MourãoNoch keine Bewertungen
- Trabalho de BiologiaDokument5 SeitenTrabalho de BiologiatodopoderosoelshadaiNoch keine Bewertungen
- Apostila Propagação de FrutiferasDokument26 SeitenApostila Propagação de FrutiferasJonas CunhaNoch keine Bewertungen
- 1.1 - Ficha Informativa 1 - Reprodução AssexuadaDokument8 Seiten1.1 - Ficha Informativa 1 - Reprodução AssexuadaElisabete CortinhasNoch keine Bewertungen
- PPPPDokument23 SeitenPPPPMicaela SantosNoch keine Bewertungen
- Diversidade Dos VegetaisDokument15 SeitenDiversidade Dos VegetaisREDINEYY FERNANDES100% (1)
- O FeijãoDokument11 SeitenO FeijãoWilton DuarteNoch keine Bewertungen
- LLLLDokument15 SeitenLLLLMicaela SantosNoch keine Bewertungen
- Germinação e DormênciaDokument31 SeitenGerminação e DormênciaRenata Girão FirmoNoch keine Bewertungen
- Aula Pratica 1 - SementesDokument7 SeitenAula Pratica 1 - SementesVicentina CotaNoch keine Bewertungen
- Vascular EsDokument4 SeitenVascular EsZecametri Carlos ZaipafaneNoch keine Bewertungen
- Cap.2.Estrutura, Composição e Propriedades Dos GrãosDokument17 SeitenCap.2.Estrutura, Composição e Propriedades Dos GrãosCarla MoretiNoch keine Bewertungen
- Texto de Apoio Sobre o Reino PlantaeDokument7 SeitenTexto de Apoio Sobre o Reino Plantaehelio magueleNoch keine Bewertungen
- PortifolioDokument8 SeitenPortifoliodeise alessandraNoch keine Bewertungen
- Multiplicacao de Plantas PDFDokument155 SeitenMultiplicacao de Plantas PDFCatarina IsabelNoch keine Bewertungen
- BotânicaDokument47 SeitenBotânicamaahonlydoguiNoch keine Bewertungen
- HHHHHHDokument14 SeitenHHHHHHMicaela SantosNoch keine Bewertungen
- BRIÓFITASDokument7 SeitenBRIÓFITASLeonardo LeidensNoch keine Bewertungen
- Órgãos Vegetativos e Reprodutivos + Vamos PraticarDokument7 SeitenÓrgãos Vegetativos e Reprodutivos + Vamos PraticarHilma BorgesNoch keine Bewertungen
- Germinacão SementesDokument38 SeitenGerminacão SementesGisely FernandesNoch keine Bewertungen
- 2 Prova de BotânicaDokument3 Seiten2 Prova de BotânicaRobenilson AlvesNoch keine Bewertungen
- Germinacao e Dormencia - 1Dokument41 SeitenGerminacao e Dormencia - 1Rachel VilaniNoch keine Bewertungen
- 1 - Reino Vegetal - Raiz Caule Folhas Flores Frutos e SementesDokument86 Seiten1 - Reino Vegetal - Raiz Caule Folhas Flores Frutos e SementesSilvia Regina Meira100% (1)
- Relat BioVegetalDokument20 SeitenRelat BioVegetalsilvana_wenceslau67% (3)
- Diversidade Das PlantasDokument11 SeitenDiversidade Das PlantasMariane MassafraNoch keine Bewertungen
- Programa CiênciaDokument16 SeitenPrograma CiênciaSegismundo Santos BarbosaNoch keine Bewertungen
- Questionario de MorfologiaDokument3 SeitenQuestionario de MorfologiaPedroNoch keine Bewertungen
- Relatório Preparo de SoluçõesDokument9 SeitenRelatório Preparo de SoluçõesjvoliveerNoch keine Bewertungen
- Relatório Titulação Ácido-BaseDokument6 SeitenRelatório Titulação Ácido-BasejvoliveerNoch keine Bewertungen
- Relatorio Determinação Da Densidade de Líquidos e SólidosDokument5 SeitenRelatorio Determinação Da Densidade de Líquidos e SólidosjvoliveerNoch keine Bewertungen
- Magnoliales e LauralesDokument10 SeitenMagnoliales e LauralesjvoliveerNoch keine Bewertungen
- 2.10.COC - Metabolismo 1Dokument15 Seiten2.10.COC - Metabolismo 1Christiane BarbiNoch keine Bewertungen
- Apostila Modulo X Biossintese de CarboidratosDokument50 SeitenApostila Modulo X Biossintese de CarboidratosVinicius RiannNoch keine Bewertungen
- Metabolismo Das ProteínasDokument5 SeitenMetabolismo Das Proteínasrsgon81Noch keine Bewertungen
- Metabolismo de CarboidratosDokument4 SeitenMetabolismo de CarboidratosHellycláudia ChavesNoch keine Bewertungen
- Gliconeogenese e Metabolismo de CarboidratosDokument12 SeitenGliconeogenese e Metabolismo de CarboidratosSuellenKartalian100% (1)
- Gliconeogenese e Via Das Pentoses FosfatosDokument33 SeitenGliconeogenese e Via Das Pentoses Fosfatoscarolaine santos100% (1)
- Metabolismo Dos Carboidratos, Respiração Celular e DesnutriçãoDokument31 SeitenMetabolismo Dos Carboidratos, Respiração Celular e DesnutriçãoLaila LongenNoch keine Bewertungen
- Resposta Endócrino Metabólica Ao TraumaDokument5 SeitenResposta Endócrino Metabólica Ao TraumameddiogomeloNoch keine Bewertungen
- Estudo Dirigido Metabolismo de CarboidratosDokument6 SeitenEstudo Dirigido Metabolismo de CarboidratosThaís CorrêaNoch keine Bewertungen
- Documento Sem TítuloDokument2 SeitenDocumento Sem TítuloAnna kassiaNoch keine Bewertungen
- Ebook Jejum Intermitente Mitos e VerdadesDokument20 SeitenEbook Jejum Intermitente Mitos e VerdadesmelissalippeNoch keine Bewertungen
- Gliconeogenese e Metabolismo Do GlicogênioDokument27 SeitenGliconeogenese e Metabolismo Do GlicogênioRicardo BorgesNoch keine Bewertungen
- Gliconeogênese Seminario Rodolfo 29 de Outubro 2021.2Dokument7 SeitenGliconeogênese Seminario Rodolfo 29 de Outubro 2021.2MC CHACALNoch keine Bewertungen
- Metabolismo Dos AminoácidosDokument44 SeitenMetabolismo Dos AminoácidosEmanuelle Thayná100% (1)
- RemitDokument7 SeitenRemitdaysemedNoch keine Bewertungen
- Bioquímica-Fisiológica-Exame 210129 010644Dokument7 SeitenBioquímica-Fisiológica-Exame 210129 010644Joana Carolina Teixeira RibeiroNoch keine Bewertungen
- Prova Bioquímica ENFERMAGEMDokument6 SeitenProva Bioquímica ENFERMAGEMSLANGIENoch keine Bewertungen
- Bioquimica Aula 1Dokument64 SeitenBioquimica Aula 1samara meira100% (1)
- Bioenergética e MetabolismoDokument47 SeitenBioenergética e MetabolismoValeskaSenaNoch keine Bewertungen
- A Vaca Leiteira Do Século 21: Lições de Metabolismo e NutriçãoDokument348 SeitenA Vaca Leiteira Do Século 21: Lições de Metabolismo e Nutriçãovenom sturckimNoch keine Bewertungen
- Casos Clínicos I Diabetes MellitusDokument17 SeitenCasos Clínicos I Diabetes MellitusNadabia Alisson BatataNoch keine Bewertungen
- Nutricionista Concurseira (Aula Gratuita para Residências)Dokument12 SeitenNutricionista Concurseira (Aula Gratuita para Residências)Daniele Souza100% (2)
- Digestão e Absorção de GlicídiosDokument4 SeitenDigestão e Absorção de Glicídiosfernandoribeirojr98Noch keine Bewertungen
- Biologia Exercicios Fisiologia Animal Sistema Endocrino Gabarito ResolucaoDokument5 SeitenBiologia Exercicios Fisiologia Animal Sistema Endocrino Gabarito ResolucaoSergio AraujoNoch keine Bewertungen
- GliconeogeneseDokument4 SeitenGliconeogeneseAline PereiraNoch keine Bewertungen
- N Terapia Clark en Diabetes Tipos 1 y 2 PortuguesDokument65 SeitenN Terapia Clark en Diabetes Tipos 1 y 2 Portuguesviviane custodio100% (2)
- Integração MetabólicaDokument2 SeitenIntegração MetabólicakarineNoch keine Bewertungen
- 6 - Metabolismo de Carboidratos I - Glicólise e GliconeogêneseDokument8 Seiten6 - Metabolismo de Carboidratos I - Glicólise e GliconeogêneseFelipe RostirollaNoch keine Bewertungen
- Exercícios RO IDokument18 SeitenExercícios RO ICarol MendesNoch keine Bewertungen
- Terapia Nutricional em Paciente GraveDokument7 SeitenTerapia Nutricional em Paciente GraveAlineNoch keine Bewertungen