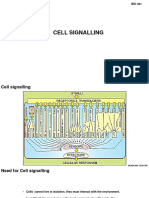
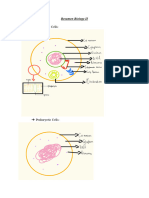
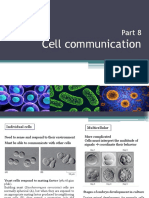
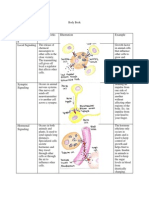
Das könnte Ihnen auch gefallen
- A-level Biology Revision: Cheeky Revision ShortcutsVon EverandA-level Biology Revision: Cheeky Revision ShortcutsBewertung: 5 von 5 Sternen5/5 (5)
- NO SignallingDokument5 SeitenNO SignallingnehashuklaNoch keine Bewertungen
- Signalling in CellsDokument11 SeitenSignalling in CellsAditi JhaNoch keine Bewertungen
- Cell Signalling and CommunicationsDokument4 SeitenCell Signalling and CommunicationsJames IbrahimNoch keine Bewertungen
- Oxygen Molecules Chemical Energy Carbon DioxideDokument3 SeitenOxygen Molecules Chemical Energy Carbon DioxideCaryl Kristen C. SulimaNoch keine Bewertungen
- Cells Typically Communicate Using Chemical SignalsDokument5 SeitenCells Typically Communicate Using Chemical SignalsrmndzrmeNoch keine Bewertungen
- UG SEM-6 (H) - CC-XIII Signal TransductionDokument22 SeitenUG SEM-6 (H) - CC-XIII Signal Transductionn7sspyywc6Noch keine Bewertungen
- Cell Signaling: Your Ad Could Be Here! Click Here To InquireDokument7 SeitenCell Signaling: Your Ad Could Be Here! Click Here To InquireRionNoch keine Bewertungen
- Introduction To Cell SignalingDokument6 SeitenIntroduction To Cell SignalingAreeba KhanNoch keine Bewertungen
- Cell Signaling: Cell Signaling Is Part of A Complex System of Communication That Governs BasicDokument6 SeitenCell Signaling: Cell Signaling Is Part of A Complex System of Communication That Governs BasicMohan RaajNoch keine Bewertungen
- Signaling Molecules and Their ReceptorsDokument13 SeitenSignaling Molecules and Their ReceptorsShweta SinghNoch keine Bewertungen
- K7 - Komunikasi Antar SelDokument30 SeitenK7 - Komunikasi Antar SelJuliana Sari HarahapNoch keine Bewertungen
- Understanding Cell Functions in The Human Body Summary PDFDokument11 SeitenUnderstanding Cell Functions in The Human Body Summary PDFcorazondelatorre12345Noch keine Bewertungen
- Chapter 15- Jnu Cellular Signaling - 複本Dokument49 SeitenChapter 15- Jnu Cellular Signaling - 複本Wai Kwong ChiuNoch keine Bewertungen
- Cell Communications and Signal Transduction: Biokimia Dan Biologi Molekuler FK UntarDokument112 SeitenCell Communications and Signal Transduction: Biokimia Dan Biologi Molekuler FK UntarAlbert TandyNoch keine Bewertungen
- Cell Signalling BasicsDokument26 SeitenCell Signalling Basics6ypywxdqmkNoch keine Bewertungen
- Cell Signaling: What Kind of Signals Do Cells Receive?Dokument5 SeitenCell Signaling: What Kind of Signals Do Cells Receive?bilalmasNoch keine Bewertungen
- Assingment Cytology-1Dokument4 SeitenAssingment Cytology-1Abdiwahab AbdirasakNoch keine Bewertungen
- CH 11 Study Guide From OnlineDokument10 SeitenCH 11 Study Guide From Onlineюрий локтионовNoch keine Bewertungen
- Cell Signaling-2021Dokument16 SeitenCell Signaling-2021VINE TUBENoch keine Bewertungen
- Tell Them What You're Going To Tell Them. Tell Them. Tell Them What You Just Told Them. AristotleDokument17 SeitenTell Them What You're Going To Tell Them. Tell Them. Tell Them What You Just Told Them. AristotleSafiya JamesNoch keine Bewertungen
- Chapter 7: Cell Communication and Signaling 1: Dr. Ivonne BejaranoDokument29 SeitenChapter 7: Cell Communication and Signaling 1: Dr. Ivonne BejaranossaNoch keine Bewertungen
- Type of Cell Junctions 1Dokument12 SeitenType of Cell Junctions 1shahrul mukminNoch keine Bewertungen
- Cell SignallingDokument32 SeitenCell SignallingMohamed NourNoch keine Bewertungen
- Cell Communication:: Signal TransductionDokument12 SeitenCell Communication:: Signal TransductionDaniloFRNoch keine Bewertungen
- Cell PhysiologyDokument7 SeitenCell PhysiologyReslie Joy Serrano100% (1)
- Kami Export - CH 11 Reading Guide PDFDokument14 SeitenKami Export - CH 11 Reading Guide PDFJinal Patel100% (3)
- Activity 1 (Physiology)Dokument4 SeitenActivity 1 (Physiology)dianarosedejorasNoch keine Bewertungen
- Biochemistry - Signal TransductionDokument4 SeitenBiochemistry - Signal Transductionsebasmarq2001Noch keine Bewertungen
- Resumen Biology II Células y Sus Partes en InglésDokument2 SeitenResumen Biology II Células y Sus Partes en Inglésmicaalvarez3001Noch keine Bewertungen
- Endocrine Physiol LectDokument22 SeitenEndocrine Physiol LectVivek ChaudharyNoch keine Bewertungen
- Part 8 Cell CommunicationDokument28 SeitenPart 8 Cell CommunicationLê Thanh HằngNoch keine Bewertungen
- Explanations For Major Oral Exam PhysioDokument5 SeitenExplanations For Major Oral Exam PhysioJolly S. SendinNoch keine Bewertungen
- Cell-Cell Interactions: Concept OutlineDokument18 SeitenCell-Cell Interactions: Concept Outlineসোমনাথ মহাপাত্রNoch keine Bewertungen
- Blok 3 (Pensinyalan Sel)Dokument8 SeitenBlok 3 (Pensinyalan Sel)cresentiaNoch keine Bewertungen
- DR Norhan PDF Lecture 3 Signal Transduction For Students Elalamein 2021-22Dokument4 SeitenDR Norhan PDF Lecture 3 Signal Transduction For Students Elalamein 2021-22Ahmed MagdyNoch keine Bewertungen
- Cell SignalingDokument20 SeitenCell Signalingnarehan13Noch keine Bewertungen
- Cell Communication and TransportDokument65 SeitenCell Communication and TransportCecilia MukototsiNoch keine Bewertungen
- Describe The Functions of The Circulatory System. (5 Marks)Dokument5 SeitenDescribe The Functions of The Circulatory System. (5 Marks)MARYAM AISYA ABDULLAH100% (1)
- Tissue Repair: Cell Regeneration and FibrosisDokument38 SeitenTissue Repair: Cell Regeneration and Fibrosisناصر دويكاتNoch keine Bewertungen
- Unit 3 Notes - Communitcation Integration and HomeostasisDokument46 SeitenUnit 3 Notes - Communitcation Integration and HomeostasisValentina CaravelisNoch keine Bewertungen
- Cell Communication PDFDokument23 SeitenCell Communication PDFSacharissa DavitaNoch keine Bewertungen
- Mekanisme Kerja Hormon Dan ReseptorDokument44 SeitenMekanisme Kerja Hormon Dan ReseptorOriza Rifki RamadanNoch keine Bewertungen
- Unit 1Dokument8 SeitenUnit 1Azeem FaizalNoch keine Bewertungen
- Saqs - (Very Important)Dokument67 SeitenSaqs - (Very Important)BRIGHTON JOSHUANoch keine Bewertungen
- Signaling Pathways PowerpointDokument36 SeitenSignaling Pathways PowerpointdiahNoch keine Bewertungen
- AP Biology Exam Review QuestionsDokument17 SeitenAP Biology Exam Review Questionsdstaples7100% (10)
- Define Receptors and Explain Signal Transduction Mechanism For Plasma Membrane ReceptorsDokument8 SeitenDefine Receptors and Explain Signal Transduction Mechanism For Plasma Membrane ReceptorsArslan Muhammad EjazNoch keine Bewertungen
- 11 DetailLectOutDokument13 Seiten11 DetailLectOuthaha_le12Noch keine Bewertungen
- 1.basic Principles of Human PhysiologyDokument94 Seiten1.basic Principles of Human PhysiologyBobyAnugrah50% (2)
- Cell Signalling in PlantsDokument51 SeitenCell Signalling in PlantsAdwika DeoNoch keine Bewertungen
- Biology SimplifiedDokument59 SeitenBiology Simplifiedoussama6928Noch keine Bewertungen
- Cell Bio AssignmentDokument4 SeitenCell Bio AssignmentHumma NayyarNoch keine Bewertungen
- Cell SignallingDokument19 SeitenCell SignallingMildred Ibe CabulisanNoch keine Bewertungen
- Cell-Cell Signaling - Masika 2021Dokument50 SeitenCell-Cell Signaling - Masika 2021omarswaleh490Noch keine Bewertungen
- AP Bio Ch. 11 Cell CommunicationDokument78 SeitenAP Bio Ch. 11 Cell CommunicationRavider JadejaNoch keine Bewertungen
- AssignmentLec2 DimnatangDokument2 SeitenAssignmentLec2 DimnatangFatimah DimnatangNoch keine Bewertungen
- AP BIOLOGY Unit 5 Cell Signaling Review Packet Answer Key-1Dokument5 SeitenAP BIOLOGY Unit 5 Cell Signaling Review Packet Answer Key-1Jeankarlo ReiNoch keine Bewertungen
- Cell Communicatio N How It Works Illustration ExampleDokument8 SeitenCell Communicatio N How It Works Illustration ExampleMaha HusseiniNoch keine Bewertungen
- Cellular Signalling SAC ChecklistDokument13 SeitenCellular Signalling SAC ChecklistqwertyuiopNoch keine Bewertungen
- Vet Pathol 2011 2011 ACVP Annual Meeting E1 E51Dokument52 SeitenVet Pathol 2011 2011 ACVP Annual Meeting E1 E51Haroon RashidNoch keine Bewertungen
- Samples of Persuasive EssaysDokument8 SeitenSamples of Persuasive Essaysb72bgzr0100% (2)
- Introduction To Medicinal Chemistry 1431 PDFDokument230 SeitenIntroduction To Medicinal Chemistry 1431 PDFjames mellaleievNoch keine Bewertungen
- Cancer CellDokument140 SeitenCancer CellmanbastiNoch keine Bewertungen
- Health Technology AssessmentDokument192 SeitenHealth Technology AssessmentĐạt Trần TiếnNoch keine Bewertungen
- Breast Cancer Therapeutics and Biomarkers: Past, Present, and Future ApproachesDokument19 SeitenBreast Cancer Therapeutics and Biomarkers: Past, Present, and Future ApproachesDavide RadiceNoch keine Bewertungen
- Neurotransmitter Cell CommunicationDokument25 SeitenNeurotransmitter Cell CommunicationOtnil DNoch keine Bewertungen
- Pediatric RhabdomyosarcomaDokument24 SeitenPediatric RhabdomyosarcomaMikael AngelooNoch keine Bewertungen
- ScendgfrytzyauDokument169 SeitenScendgfrytzyaufernangomezraNoch keine Bewertungen
- Treatment of Acute Lymphoblastic Leukemia: Review ArticleDokument13 SeitenTreatment of Acute Lymphoblastic Leukemia: Review ArticlehoshihopeNoch keine Bewertungen
- Classification of OncogenesDokument20 SeitenClassification of Oncogenes18B236 - NIVYA SUBULAXMI MNoch keine Bewertungen
- Biol 309 Question Bank Cell CommunicationDokument6 SeitenBiol 309 Question Bank Cell CommunicationWalaa abo fool100% (1)
- Signals and ReceptorsDokument28 SeitenSignals and ReceptorsEmilio MurciaNoch keine Bewertungen
- Antineoplastic AgentsDokument25 SeitenAntineoplastic AgentsAlyssaGrandeMontimorNoch keine Bewertungen
- AntitumoralesDokument19 SeitenAntitumoralesMaira BianchiniNoch keine Bewertungen
- ChemotherapyDokument16 SeitenChemotherapyNandhini SaranathanNoch keine Bewertungen
- New Drugs 2018: New Drug Mechanism of Action UseDokument7 SeitenNew Drugs 2018: New Drug Mechanism of Action UsePremangshu GhoshalNoch keine Bewertungen
- Gene Mutation ReportDokument7 SeitenGene Mutation ReportArjun SanchetiNoch keine Bewertungen
- Pharmacology Quizlet: Chapter 1 Intro To PharmacologyDokument18 SeitenPharmacology Quizlet: Chapter 1 Intro To PharmacologyJerome ValerianoNoch keine Bewertungen
- Jie Jack Li - Medicinal Chemistry For Practitioners-John Wiley & Sons Inc (2021)Dokument401 SeitenJie Jack Li - Medicinal Chemistry For Practitioners-John Wiley & Sons Inc (2021)Rotoplastic de Cuauhtémoc S.A. de C.V.100% (1)
- RoscvitineDokument99 SeitenRoscvitineTommyNoch keine Bewertungen
- High Throughput Screening - Compound Library - Chemical Library - Screening LibraryDokument12 SeitenHigh Throughput Screening - Compound Library - Chemical Library - Screening LibrarySelleck Chemicals LLCNoch keine Bewertungen
- Signal Trans Duct IonDokument13 SeitenSignal Trans Duct IonPrasath ThevarajahNoch keine Bewertungen
- Cancer ChemotherapyDokument37 SeitenCancer ChemotherapyRubyrose TagumNoch keine Bewertungen
- Learning Outcome.: Terms To Know/keywordsDokument5 SeitenLearning Outcome.: Terms To Know/keywordsiknowu250Noch keine Bewertungen
- Oncogenes and Tumour Suppressor GenesDokument59 SeitenOncogenes and Tumour Suppressor GenesXutjie FairdianaNoch keine Bewertungen
- Enzyme Linked Receptors LectureDokument52 SeitenEnzyme Linked Receptors LectureJason PastisNoch keine Bewertungen
- Chapter 18 Test Bank PatrickDokument65 SeitenChapter 18 Test Bank PatrickAmir ali WalizadehNoch keine Bewertungen
- Lecture 2 Membrane Transport & Cell Signaling - QuestionsDokument6 SeitenLecture 2 Membrane Transport & Cell Signaling - Questions中华雅思王Noch keine Bewertungen
- Self - Sufficiency in Growth Signals Cancer.Dokument21 SeitenSelf - Sufficiency in Growth Signals Cancer.siddhant kushareNoch keine Bewertungen