Das könnte Ihnen auch gefallen
- Hällbrink Et Al. - 2001 - Cargo Delivery Kinetics of Cell-Penetrating PeptidDokument9 SeitenHällbrink Et Al. - 2001 - Cargo Delivery Kinetics of Cell-Penetrating Peptidquentin pNoch keine Bewertungen
- Targeting Calcium Signaling in Cancer TherapyDokument15 SeitenTargeting Calcium Signaling in Cancer TherapyLeonel LedezmaNoch keine Bewertungen
- ''Third Messengers''? Extracellular Calcium and cAMP: Second Messengers AsDokument9 Seiten''Third Messengers''? Extracellular Calcium and cAMP: Second Messengers Asanusha jrNoch keine Bewertungen
- Pathological and Chemical Effectors of The Erythrocyte Calcium Pumping Protein: A Review.Dokument15 SeitenPathological and Chemical Effectors of The Erythrocyte Calcium Pumping Protein: A Review.Clement BewajiNoch keine Bewertungen
- Investigating Transport Proteins by Solid State NMRDokument14 SeitenInvestigating Transport Proteins by Solid State NMRAArriiss WizushkiNoch keine Bewertungen
- Cancer Resistance 2Dokument10 SeitenCancer Resistance 2Gregory LaskarisNoch keine Bewertungen
- Cornichon 2Dokument8 SeitenCornichon 2bryanmendietaNoch keine Bewertungen
- Please Use Endnote Program To Organize Your ReferencesDokument25 SeitenPlease Use Endnote Program To Organize Your ReferencesMy PlanNoch keine Bewertungen
- Soysa 2013Dokument12 SeitenSoysa 2013E May RamNoch keine Bewertungen
- Aps2008132 PDFDokument9 SeitenAps2008132 PDFSumant SainiNoch keine Bewertungen
- Lecture 5 Signal Transduction and Cellular ResponseDokument52 SeitenLecture 5 Signal Transduction and Cellular Responseapi-251915360Noch keine Bewertungen
- European Journal of Biochemistry - 2001 - Kalies - Protein Translocation Into The Endoplasmic Reticulum ERDokument5 SeitenEuropean Journal of Biochemistry - 2001 - Kalies - Protein Translocation Into The Endoplasmic Reticulum ER12 yashika palNoch keine Bewertungen
- Erythrocyte Membrane-Camouflaged Polymeric Nanoparticles As A Biomimetic Delivery PlatformDokument6 SeitenErythrocyte Membrane-Camouflaged Polymeric Nanoparticles As A Biomimetic Delivery PlatformHyukLadezaNoch keine Bewertungen
- AnimalPhysiology - Membrane Structure and FunctionDokument34 SeitenAnimalPhysiology - Membrane Structure and FunctionsantoshNoch keine Bewertungen
- The Structural Basis of Transcription: 10 Years After The Nobel Prize in ChemistryDokument10 SeitenThe Structural Basis of Transcription: 10 Years After The Nobel Prize in ChemistrySergio Davila GonzalesNoch keine Bewertungen
- Olson-Sahai2008 Article TheActinCytoskeletonInCancerCeDokument15 SeitenOlson-Sahai2008 Article TheActinCytoskeletonInCancerCeFernanda LópezNoch keine Bewertungen
- Calcium Signaling: Encoding and Decoding: Advanced ArticleDokument10 SeitenCalcium Signaling: Encoding and Decoding: Advanced ArticleazzaassNoch keine Bewertungen
- Nuclear Trafficking in Health and Disease: SciencedirectDokument8 SeitenNuclear Trafficking in Health and Disease: SciencedirectMariaNoch keine Bewertungen
- How Viruses Access The NucleusDokument12 SeitenHow Viruses Access The NucleusGABRIEL GIANASMIDISNoch keine Bewertungen
- Peinado Ca-Photorelease PNAS2015Dokument9 SeitenPeinado Ca-Photorelease PNAS2015Juan David Avila RinconNoch keine Bewertungen
- The Human Plasma ProteomeDokument23 SeitenThe Human Plasma ProteomeKarina CordovaNoch keine Bewertungen
- Aquaporin Biology and Nervous System: Buffoli BarbaraDokument8 SeitenAquaporin Biology and Nervous System: Buffoli BarbaraKhaled Abdel-saterNoch keine Bewertungen
- Bba - Biomembranes: SciencedirectDokument12 SeitenBba - Biomembranes: SciencedirectDIEGO FERNANDO TULCAN SILVANoch keine Bewertungen
- David H. J. Bunka and Peter G. Stockley - Aptamers Come of Age - at LastDokument9 SeitenDavid H. J. Bunka and Peter G. Stockley - Aptamers Come of Age - at LastGmso30% (1)
- Paper No and Title 15: Bioinorganic Chemistry Module No and Title 8: Ion Transport Through Cell Membrane and Na /K Pump Module Tag CHE - P15 - M8Dokument9 SeitenPaper No and Title 15: Bioinorganic Chemistry Module No and Title 8: Ion Transport Through Cell Membrane and Na /K Pump Module Tag CHE - P15 - M8kottooranjohnbNoch keine Bewertungen
- 2009 AemDokument8 Seiten2009 AemAndrew SingerNoch keine Bewertungen
- Calcium Signaling and Cytotoxicity: in MammalianDokument11 SeitenCalcium Signaling and Cytotoxicity: in MammalianRoland ToroNoch keine Bewertungen
- EC05 Cell Surface To Phagocytosis TextDokument8 SeitenEC05 Cell Surface To Phagocytosis TextSilvito94Noch keine Bewertungen
- Anti-Calmodulins and Tricyclic Adjuvants in Pain Therapy Block The TRPV1 ChannelDokument12 SeitenAnti-Calmodulins and Tricyclic Adjuvants in Pain Therapy Block The TRPV1 ChannelplosfanNoch keine Bewertungen
- Membrane ProteinsDokument32 SeitenMembrane ProteinsDevanand T Santha100% (1)
- Protein TargetingDokument7 SeitenProtein TargetingRaven JohnsonNoch keine Bewertungen
- Proteasome Phase Separation A Novel Layer of Quality ControlDokument2 SeitenProteasome Phase Separation A Novel Layer of Quality ControlDoniNoch keine Bewertungen
- Solution BiotechDokument54 SeitenSolution Biotechkavita sahaiNoch keine Bewertungen
- Tutorial 2, Plasma MembraneDokument51 SeitenTutorial 2, Plasma MembranesureshbabubnysNoch keine Bewertungen
- Curt Hoys 2013Dokument12 SeitenCurt Hoys 2013broken22Noch keine Bewertungen
- RBC MembraneDokument41 SeitenRBC MembranemwambungupNoch keine Bewertungen
- 2017-07 Lipid Nanoparticle Systems For Enabling Gene TherapiesDokument9 Seiten2017-07 Lipid Nanoparticle Systems For Enabling Gene TherapiesRetro ActiveNoch keine Bewertungen
- William James - AptamersDokument25 SeitenWilliam James - AptamersGmso3Noch keine Bewertungen
- BiologyDokument25 SeitenBiologyIce BearNoch keine Bewertungen
- Pahala 1Dokument5 SeitenPahala 1Daeng MangkaNoch keine Bewertungen
- Genes Dev.-2001-Abraham-2177-96Dokument21 SeitenGenes Dev.-2001-Abraham-2177-96M Faizal RezhaNoch keine Bewertungen
- Second Messenger System in Cell SignallingDokument14 SeitenSecond Messenger System in Cell SignallingMamata MishraNoch keine Bewertungen
- 2018 Lecture12Dokument31 Seiten2018 Lecture12Adam Bryant PoonawalaNoch keine Bewertungen
- CHAPTER 1 Cellular Physiology: Volume and Composition of Body Fluids, 1Dokument13 SeitenCHAPTER 1 Cellular Physiology: Volume and Composition of Body Fluids, 1mr nice guyNoch keine Bewertungen
- Nihms96320 Slo3 SpermDokument12 SeitenNihms96320 Slo3 SpermBiodiesel NegAceNoch keine Bewertungen
- Li Weber2013Dokument9 SeitenLi Weber2013Hector Javier BurgosNoch keine Bewertungen
- CHE 320 - Journal 2Dokument7 SeitenCHE 320 - Journal 2Garcia RaphNoch keine Bewertungen
- Paper 12Dokument12 SeitenPaper 12Nexi anessaNoch keine Bewertungen
- Cellpenetrating Peptides As A Novel Transdermal Drug Delivery SystemDokument8 SeitenCellpenetrating Peptides As A Novel Transdermal Drug Delivery SystemHinal AmbasanaNoch keine Bewertungen
- Special Articles Serum Albumin: Marcus Murray Oratz Sidney S. SchreiberDokument17 SeitenSpecial Articles Serum Albumin: Marcus Murray Oratz Sidney S. SchreiberTuấn AnhNoch keine Bewertungen
- The Molecular Features of Uncoupling Protein 1 Support A Conventional Mitochondrial Carrier-Like MechanismDokument36 SeitenThe Molecular Features of Uncoupling Protein 1 Support A Conventional Mitochondrial Carrier-Like Mechanismpasindu chamodNoch keine Bewertungen
- Immunological Cross-Reactivity Between Calcium ATPases From Human and Porcine ErythrocytesDokument7 SeitenImmunological Cross-Reactivity Between Calcium ATPases From Human and Porcine ErythrocytesClement BewajiNoch keine Bewertungen
- Erythrocytes-Camouflaged Nanoparticles A Promising Delivery System For Drugs and VaccinesDokument18 SeitenErythrocytes-Camouflaged Nanoparticles A Promising Delivery System For Drugs and VaccinesWang JianweiNoch keine Bewertungen
- Molecular Cell Biology Lodish 7th Edition Solutions ManualDokument8 SeitenMolecular Cell Biology Lodish 7th Edition Solutions ManualKennethHendersonyecg100% (40)
- Solution Manual For Molecular Cell Biology 8th Edition Harvey Lodish Arnold Berk Chris A Kaiser Monty Krieger Anthony Bretscher Hidde Ploegh Angelika Amon Kelsey C Martin 33Dokument5 SeitenSolution Manual For Molecular Cell Biology 8th Edition Harvey Lodish Arnold Berk Chris A Kaiser Monty Krieger Anthony Bretscher Hidde Ploegh Angelika Amon Kelsey C Martin 33PhilipWoodpsen100% (39)
- The Role of Pannexin-1 Channel - ReviewDokument4 SeitenThe Role of Pannexin-1 Channel - ReviewInternational Journal of Innovative Science and Research TechnologyNoch keine Bewertungen
- Atom Transfer Radical Polymerization and The Synthesis of Polymeric MaterialsDokument15 SeitenAtom Transfer Radical Polymerization and The Synthesis of Polymeric MaterialsRawan AbdullahNoch keine Bewertungen
- 50 DiazRuiz08 PDFDokument8 Seiten50 DiazRuiz08 PDFCarolina RicárdezNoch keine Bewertungen
- PAKs, RAC/CDC42 (p21)-activated Kinases: Towards the Cure of Cancer and Other PAK-dependent DiseasesVon EverandPAKs, RAC/CDC42 (p21)-activated Kinases: Towards the Cure of Cancer and Other PAK-dependent DiseasesHiroshi MarutaNoch keine Bewertungen
- Ion Transport in ProkaryotesVon EverandIon Transport in ProkaryotesBarry P. RosenNoch keine Bewertungen
- Uji Normalitas Dan HomogenitasDokument3 SeitenUji Normalitas Dan HomogenitasiraNoch keine Bewertungen
- Laporan Praktikum Formulasi Dan Teknologi Sediaan Solida Formulasi Tablet Ibuprofen Metode Granulasi BasahDokument5 SeitenLaporan Praktikum Formulasi Dan Teknologi Sediaan Solida Formulasi Tablet Ibuprofen Metode Granulasi BasahiraNoch keine Bewertungen
- Profil Disolusi Tab AcetosalDokument1 SeiteProfil Disolusi Tab AcetosaliraNoch keine Bewertungen
- Dapus Jurnal JaheDokument2 SeitenDapus Jurnal JaheiraNoch keine Bewertungen
- Ovulation Test: 5 Important Points You Need To ReadDokument2 SeitenOvulation Test: 5 Important Points You Need To ReadiraNoch keine Bewertungen
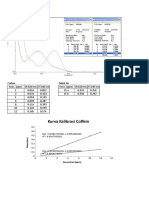
- Kurva Kalibrasi Coffein: 0.6 0.7 F (X) 0.0481373938x + 0.0052988669 R 0.9987395268Dokument1 SeiteKurva Kalibrasi Coffein: 0.6 0.7 F (X) 0.0481373938x + 0.0052988669 R 0.9987395268iraNoch keine Bewertungen
- Chart Title: 0.800 0.900 F (X) 0.0568428571x + 0.0039142857 R 0.9983149176Dokument5 SeitenChart Title: 0.800 0.900 F (X) 0.0568428571x + 0.0039142857 R 0.9983149176iraNoch keine Bewertungen
- Gambar Alat DisolusiDokument2 SeitenGambar Alat DisolusiiraNoch keine Bewertungen
- 2.1 The Nuclear AtomDokument14 Seiten2.1 The Nuclear AtomCrystalNoch keine Bewertungen
- Compressor Technical Data: NJ9232GK 220-240 V 50 HZ 943NA01 A - Application / Limit Working ConditionsDokument3 SeitenCompressor Technical Data: NJ9232GK 220-240 V 50 HZ 943NA01 A - Application / Limit Working ConditionsGeri SulanjakuNoch keine Bewertungen
- SCC 5Dokument1 SeiteSCC 5TasmanijskaNemaNoch keine Bewertungen
- Experimental Investigation On Hybrid Fibre Reinforced Concrete by Partial Replacement of MDokument33 SeitenExperimental Investigation On Hybrid Fibre Reinforced Concrete by Partial Replacement of MAarthiNoch keine Bewertungen
- 1 s2.0 S2090123213000969 MainDokument17 Seiten1 s2.0 S2090123213000969 Mainmarius_alex_32Noch keine Bewertungen
- Project File LawsDokument5 SeitenProject File LawsSonaNoch keine Bewertungen
- Birla Instritute of Technology & Science, Pilani, Pilani Campus Chemical Engineering DepartmentDokument2 SeitenBirla Instritute of Technology & Science, Pilani, Pilani Campus Chemical Engineering DepartmentVaibhav SharmaNoch keine Bewertungen
- Response of First OrderDokument4 SeitenResponse of First Order阿尔坎塔拉约翰·肯尼斯Noch keine Bewertungen
- RW Ti Pds Prorox WM 960 Astm IndiaDokument1 SeiteRW Ti Pds Prorox WM 960 Astm IndiaLee Kok RoyNoch keine Bewertungen
- Paper 6Dokument14 SeitenPaper 6UmaibalanNoch keine Bewertungen
- Tds-Antifoam Ew Defoamer Saw 2510Dokument1 SeiteTds-Antifoam Ew Defoamer Saw 2510kokotampanNoch keine Bewertungen
- Vinjey Software ShortlistDokument3 SeitenVinjey Software Shortlistnjelp2002.meNoch keine Bewertungen
- Gibbs Free Energy of Formation - Gaussian PDFDokument19 SeitenGibbs Free Energy of Formation - Gaussian PDFRudolf KiraljNoch keine Bewertungen
- Fe2O3 SourceDokument5 SeitenFe2O3 SourceZahid FarooqNoch keine Bewertungen
- Atomic Absorption Spectroscopy: Tutorial 2Dokument9 SeitenAtomic Absorption Spectroscopy: Tutorial 2MSKNoch keine Bewertungen
- Physical Science - Week 28Dokument4 SeitenPhysical Science - Week 28Mira VeranoNoch keine Bewertungen
- TECKA Air Cooled Screw Chiller (Heat Pump) Unit - Brochure TECKA202103Dokument10 SeitenTECKA Air Cooled Screw Chiller (Heat Pump) Unit - Brochure TECKA202103Guvanch MuradowNoch keine Bewertungen
- 2012 Env Drill Comp Fluids Dir PDFDokument25 Seiten2012 Env Drill Comp Fluids Dir PDFWilson WanNoch keine Bewertungen
- Atmospheric AerosolsDokument24 SeitenAtmospheric Aerosolsh_guthrieNoch keine Bewertungen
- MUCLecture 2023 3140165Dokument9 SeitenMUCLecture 2023 3140165hus khNoch keine Bewertungen
- TOS-G8 FORCE MOTION AND ENERGY (1st Grading) .Docx Version 1Dokument6 SeitenTOS-G8 FORCE MOTION AND ENERGY (1st Grading) .Docx Version 1Kimberly Sumbillo MonticalboNoch keine Bewertungen
- SmartScan Manual Issue 2.5Dokument39 SeitenSmartScan Manual Issue 2.5Nayana SareenNoch keine Bewertungen
- 06 Lecture 6 CES Edupack ProgrammeDokument12 Seiten06 Lecture 6 CES Edupack Programmeaaroncete14Noch keine Bewertungen
- Neutrinos in Cosmology - A. DolgovDokument379 SeitenNeutrinos in Cosmology - A. DolgovR. Gray100% (1)
- To Study Rate of DiffusionDokument17 SeitenTo Study Rate of DiffusionShlok Singh100% (1)
- Enhancing SF6 Analysis - CIGREDokument58 SeitenEnhancing SF6 Analysis - CIGREJoseph SumpaicoNoch keine Bewertungen
- Loctite 5182Dokument3 SeitenLoctite 5182ccorp0089Noch keine Bewertungen
- Chapter+4+ +Imperfections+in+CrystalsDokument25 SeitenChapter+4+ +Imperfections+in+Crystalspramod_kNoch keine Bewertungen
- Wrought 18Chromium-14Nickel-2.5Molybdenum Stainless Steel Sheet and Strip For Surgical Implants (UNS S31673)Dokument4 SeitenWrought 18Chromium-14Nickel-2.5Molybdenum Stainless Steel Sheet and Strip For Surgical Implants (UNS S31673)Angel ChangNoch keine Bewertungen
- Article 5.0: Hazardous (Classified) Locations, Classes I, Ii, and Iii, Divisions 1 and 2Dokument22 SeitenArticle 5.0: Hazardous (Classified) Locations, Classes I, Ii, and Iii, Divisions 1 and 2RodrinerBillonesNoch keine Bewertungen