Das könnte Ihnen auch gefallen
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
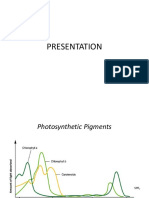
- PHOTOSYNTHESIS Lab ActivityDokument4 SeitenPHOTOSYNTHESIS Lab ActivityMaybelynNoch keine Bewertungen
- Grade 6 English Reading Identifying Sub Heading For A Given HeadingDokument6 SeitenGrade 6 English Reading Identifying Sub Heading For A Given HeadingJuan Alas Ronaldo AziongNoch keine Bewertungen
- M2 - General Bio 1 - Q2Dokument10 SeitenM2 - General Bio 1 - Q2Remilyn GonzalesNoch keine Bewertungen
- Effect of Calcium Nitrate and Potassium Nitrate Priming On Seed Germination and Seedling Vigour of Papaya (Carica Papaya L.)Dokument6 SeitenEffect of Calcium Nitrate and Potassium Nitrate Priming On Seed Germination and Seedling Vigour of Papaya (Carica Papaya L.)boonyongchiraNoch keine Bewertungen
- High Throughput Analysis of Leaf Chlorophyll Content in Sorghum Using RGB, Hyperspectral, and Fluorescence Imaging and Sensor FusionDokument17 SeitenHigh Throughput Analysis of Leaf Chlorophyll Content in Sorghum Using RGB, Hyperspectral, and Fluorescence Imaging and Sensor FusionIván EstevesNoch keine Bewertungen
- Nabila Tisa Lathifah - Week 2Dokument11 SeitenNabila Tisa Lathifah - Week 2Nabila LathifahNoch keine Bewertungen
- 1 - Science Reading ComprehensionDokument10 Seiten1 - Science Reading ComprehensionFatihUyakNoch keine Bewertungen
- Photosynthesis in Higher Plants: NEET 2022 Group E and ADokument14 SeitenPhotosynthesis in Higher Plants: NEET 2022 Group E and AShruti KumariNoch keine Bewertungen
- Smart Nano Particles TechnologyDokument588 SeitenSmart Nano Particles TechnologyJosé RamírezNoch keine Bewertungen
- Cyanobacteria As Photoautotrophic BiofactoriesDokument32 SeitenCyanobacteria As Photoautotrophic BiofactoriesAbel YapoNoch keine Bewertungen
- IAL Biology SB2 AnswersDokument77 SeitenIAL Biology SB2 AnswersChryssa Economou100% (7)
- 1 PBDokument8 Seiten1 PBVelani ElendiaNoch keine Bewertungen
- Biology Lab 5 FinalDokument5 SeitenBiology Lab 5 FinalAlice Lin75% (12)
- Aging of PlantDokument30 SeitenAging of PlantMujahid HussainNoch keine Bewertungen
- 10 Dangerous Toothpaste IngredientsDokument13 Seiten10 Dangerous Toothpaste IngredientsKilaru Harsha VardhanNoch keine Bewertungen
- Gen Bio 1 - Quarter 2 Summative Test 1Dokument2 SeitenGen Bio 1 - Quarter 2 Summative Test 1Syrine Myles Sullivan100% (2)
- Gen Bio ReviewerDokument4 SeitenGen Bio ReviewerhahatdogNoch keine Bewertungen
- D 3731 - 87 R04 - Rdm3mzeDokument4 SeitenD 3731 - 87 R04 - Rdm3mzeAifam RawNoch keine Bewertungen
- Photosynthesis Summary ParagraphDokument9 SeitenPhotosynthesis Summary ParagraphStephen MceleaveyNoch keine Bewertungen
- PresentationDokument16 SeitenPresentationASIF ADNANNoch keine Bewertungen
- Brochure Dualex 1Dokument6 SeitenBrochure Dualex 1Manish GargNoch keine Bewertungen
- General Biology 1: Quarter 2 - Module 1: ATP-ADP CycleDokument26 SeitenGeneral Biology 1: Quarter 2 - Module 1: ATP-ADP CyclePLS help me get 1k subsNoch keine Bewertungen
- 4th Olympiad of Metropolises Chemistry: September 4, 2019 Moscow, RussiaDokument23 Seiten4th Olympiad of Metropolises Chemistry: September 4, 2019 Moscow, Russiamila_a_vaNoch keine Bewertungen
- Understanding of The: Science Teacher's GuideDokument2 SeitenUnderstanding of The: Science Teacher's GuideEldie OcarizaNoch keine Bewertungen
- Mapa de Vegetación Cd. Júarez ChihuahuaDokument4 SeitenMapa de Vegetación Cd. Júarez ChihuahuaMi Si HoNoch keine Bewertungen
- Effects of Salinity and Nutrients On The Growth and Chlorophyll Uorescence of Caulerpa LentilliferaDokument10 SeitenEffects of Salinity and Nutrients On The Growth and Chlorophyll Uorescence of Caulerpa LentilliferaDadi KamaludinNoch keine Bewertungen
- Standard Operating Procedures: Chlorophyll DeterminationDokument6 SeitenStandard Operating Procedures: Chlorophyll DeterminationMAYUR CHATURNoch keine Bewertungen
- HW9 SolutionsDokument5 SeitenHW9 Solutionskakaish100Noch keine Bewertungen
- Chlorophyll A Determination by FluorescenceDokument22 SeitenChlorophyll A Determination by FluorescencefrfunkNoch keine Bewertungen
- Plant NutritionDokument155 SeitenPlant NutritionRadu100% (1)