Das könnte Ihnen auch gefallen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Chapter 03Dokument75 SeitenChapter 03Tony DiPierryNoch keine Bewertungen
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Seeley's Chapter 8 Nervous SystemDokument100 SeitenSeeley's Chapter 8 Nervous SystemChristine Tapawan88% (8)
- Hand Function in The ChildDokument476 SeitenHand Function in The ChildOmar Najera de GonzalezNoch keine Bewertungen
- Surgery Illustrated - Focus - Tapering of The MegaureterDokument5 SeitenSurgery Illustrated - Focus - Tapering of The MegauretermoetazNoch keine Bewertungen
- The Prostate ExamDokument13 SeitenThe Prostate ExammoetazNoch keine Bewertungen
- Clinical Anatomy of Anterior Abdominal Wall - MEDCHROMEDokument4 SeitenClinical Anatomy of Anterior Abdominal Wall - MEDCHROMEmoetazNoch keine Bewertungen
- Uro Embryology Cunha 2004Dokument16 SeitenUro Embryology Cunha 2004moetazNoch keine Bewertungen
- Development of AUA Clinical Practice GuidelinesDokument14 SeitenDevelopment of AUA Clinical Practice GuidelinesmoetazNoch keine Bewertungen
- IPSS QuizDokument2 SeitenIPSS QuizmoetazNoch keine Bewertungen
- Anatomy of The Anterior Abdominal Wall & Groin PDFDokument4 SeitenAnatomy of The Anterior Abdominal Wall & Groin PDFmoetazNoch keine Bewertungen
- Rusch Urology BroschureDokument72 SeitenRusch Urology BroschuremoetazNoch keine Bewertungen
- Modified Gibson Incision - WikipediaDokument1 SeiteModified Gibson Incision - WikipediamoetazNoch keine Bewertungen
- BalanoposthitisDokument45 SeitenBalanoposthitismoetazNoch keine Bewertungen
- Electrosurgery - Principles, Biologic Effects and ResultsDokument16 SeitenElectrosurgery - Principles, Biologic Effects and ResultsmoetazNoch keine Bewertungen
- EUA Conference FormsDokument4 SeitenEUA Conference FormsmoetazNoch keine Bewertungen
- Gibson Incision - OPERATIVE RECIPES For UrologistsDokument1 SeiteGibson Incision - OPERATIVE RECIPES For UrologistsmoetazNoch keine Bewertungen
- Jurnal 1Dokument7 SeitenJurnal 1bungagobeNoch keine Bewertungen
- Nej Mic M 1308004Dokument1 SeiteNej Mic M 1308004moetazNoch keine Bewertungen
- Renal AutotransplantationDokument5 SeitenRenal AutotransplantationmoetazNoch keine Bewertungen
- A Simple Procedure For Management of Urethrocutaneous Fistulas Post-Hypospadias Repair - S Mohamed, N Mohamed, T Esmael, SH Khaled, African Journal of Paediatric Surgery (AJPS)Dokument3 SeitenA Simple Procedure For Management of Urethrocutaneous Fistulas Post-Hypospadias Repair - S Mohamed, N Mohamed, T Esmael, SH Khaled, African Journal of Paediatric Surgery (AJPS)moetazNoch keine Bewertungen
- Cystography 2Dokument41 SeitenCystography 2moetazNoch keine Bewertungen
- Prostate BiopsyDokument6 SeitenProstate BiopsyUr MyNoch keine Bewertungen
- (5) (1)Dokument12 Seiten(5) (1)moetazNoch keine Bewertungen
- MCQ Uro1Dokument1 SeiteMCQ Uro1Kamal El-Ghorory67% (3)
- AUAAnatomy Small00center10 2173856Dokument5 SeitenAUAAnatomy Small00center10 2173856moetazNoch keine Bewertungen
- Post Graduate 2013Dokument4 SeitenPost Graduate 2013moetazNoch keine Bewertungen
- Cadmium in VXDokument12 SeitenCadmium in VXmoetazNoch keine Bewertungen
- Appendices PDFDokument105 SeitenAppendices PDFmoetazNoch keine Bewertungen
- Active Surveillance For Prostate Cancer AUA Update 33 2008Dokument8 SeitenActive Surveillance For Prostate Cancer AUA Update 33 2008moetazNoch keine Bewertungen
- Asymptomatic Microscopic HematuriaDokument10 SeitenAsymptomatic Microscopic HematuriaCarlos Felipe Campo BetancourthNoch keine Bewertungen
- AUA Adult Urodynamic Guidelines PDFDokument10 SeitenAUA Adult Urodynamic Guidelines PDFmoetaz0% (1)
- Urinary Diversion Following Cystectomy: EAU Update SeriesDokument9 SeitenUrinary Diversion Following Cystectomy: EAU Update SeriesmoetazNoch keine Bewertungen
- 7.reflex Regulation of Physiological Function PDFDokument2 Seiten7.reflex Regulation of Physiological Function PDFNektarios TsakalosNoch keine Bewertungen
- Cranial NerveshDokument1 SeiteCranial Nerveshcorreiodesportivo8071Noch keine Bewertungen
- Efferent Visual System (Ocular Motor Pathways)Dokument38 SeitenEfferent Visual System (Ocular Motor Pathways)riskhapangestika100% (1)
- Neurophysiology in Oral and Maxillofacial SurgeryDokument35 SeitenNeurophysiology in Oral and Maxillofacial SurgeryPriya ManimalaNoch keine Bewertungen
- Physiological Mechanisms of PainDokument7 SeitenPhysiological Mechanisms of PainZAFFAR QURESHINoch keine Bewertungen
- Ascending and Descending Spinal Tracts 2016Dokument20 SeitenAscending and Descending Spinal Tracts 2016Ayodeji LuckyNoch keine Bewertungen
- Brainstem Stroke SyndromesDokument43 SeitenBrainstem Stroke SyndromesSarah Choo Qian RouNoch keine Bewertungen
- Surcos MEDULA ESPINALDokument14 SeitenSurcos MEDULA ESPINALAngela MorantNoch keine Bewertungen
- Blood Supply of BrainDokument2 SeitenBlood Supply of BrainvmagtotoNoch keine Bewertungen
- Spinal CordDokument48 SeitenSpinal CordDr.Saber - Ar - Raffi100% (1)
- TrigeminalDokument12 SeitenTrigeminalÂhmed ÂmeerNoch keine Bewertungen
- Lecture - Slides-01 - 06 - Medial Surface of The BrainDokument6 SeitenLecture - Slides-01 - 06 - Medial Surface of The BrainMaria Eduarda VianaNoch keine Bewertungen
- Anat 6.6 Cerebellum - DeriadaDokument11 SeitenAnat 6.6 Cerebellum - Deriadalovelots1234Noch keine Bewertungen
- Disorders of The Nervous SystemDokument5 SeitenDisorders of The Nervous SystemAgronaSlaughterNoch keine Bewertungen
- Double Crush SyndromeDokument14 SeitenDouble Crush SyndromeJosip SulenticNoch keine Bewertungen
- Basal Ganglia Physiological AspectsDokument54 SeitenBasal Ganglia Physiological AspectsPhysiology by Dr Raghuveer100% (1)
- PSYC 21 2022 ANS Additional Notes RevisedDokument1 SeitePSYC 21 2022 ANS Additional Notes RevisedsdahjdhasjkdhaskdNoch keine Bewertungen
- Ramsay Hunt SyndromeDokument6 SeitenRamsay Hunt SyndromeSonya Ananda Ellya JohnNoch keine Bewertungen
- General Somatic Afferents: Luis Emmanuel O. Esguerra, MDDokument65 SeitenGeneral Somatic Afferents: Luis Emmanuel O. Esguerra, MDlovelots1234Noch keine Bewertungen
- NeurophysiologyDokument3 SeitenNeurophysiologydoyoung kimNoch keine Bewertungen
- Dance Anatomy Chapter 1 SummaryDokument3 SeitenDance Anatomy Chapter 1 SummaryLucia Natalia Chavez (Student)Noch keine Bewertungen
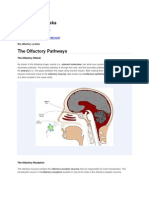
- The Olfactory PathwaysDokument3 SeitenThe Olfactory PathwaysLeo TolentinoNoch keine Bewertungen
- Human Physic Human Physic Bio Chem Test Ready Stuff of TestDokument36 SeitenHuman Physic Human Physic Bio Chem Test Ready Stuff of TestnaifNoch keine Bewertungen
- Limbic SystemDokument4 SeitenLimbic SystemAyu NurdiyanNoch keine Bewertungen
- Nervous SystemDokument2 SeitenNervous SystemShanel Aubrey AglibutNoch keine Bewertungen
- MCQ 23 Physio AnswersDokument8 SeitenMCQ 23 Physio AnswersJulyhathul Kuraishi100% (1)
- Neurology and Special Senses: High-Yield SystemsDokument72 SeitenNeurology and Special Senses: High-Yield SystemsMahmoud Abu MayalehNoch keine Bewertungen