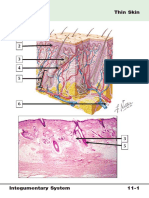
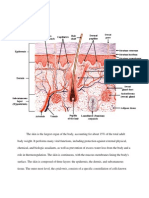
Das könnte Ihnen auch gefallen
- Human Chromosomes: An Illustrated Introduction to Human CytogeneticsVon EverandHuman Chromosomes: An Illustrated Introduction to Human CytogeneticsBewertung: 5 von 5 Sternen5/5 (1)
- The Desmosome: Cell Science Lessons From Human Diseases: 797 CommentaryDokument10 SeitenThe Desmosome: Cell Science Lessons From Human Diseases: 797 CommentaryKatja BadanjakNoch keine Bewertungen
- Number 1 Epithelial Biology: Mucosal Diseases SeriesDokument14 SeitenNumber 1 Epithelial Biology: Mucosal Diseases SeriesMoni SantamariaNoch keine Bewertungen
- Chapter 15:: Epidermal and Dermal Adhesion:: Leena Bruckner-Tuderman &Dokument7 SeitenChapter 15:: Epidermal and Dermal Adhesion:: Leena Bruckner-Tuderman &nazhifah apotekerNoch keine Bewertungen
- Epithelium Surface SpecializationsDokument41 SeitenEpithelium Surface SpecializationsMitzel SapaloNoch keine Bewertungen
- Cell JunctionsDokument30 SeitenCell JunctionsNavodit GoelNoch keine Bewertungen
- Link To Publication in University of Groningen/UMCG Research DatabaseDokument49 SeitenLink To Publication in University of Groningen/UMCG Research DatabasescribdangeloNoch keine Bewertungen
- Cell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemDokument70 SeitenCell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemGovinda BiswasNoch keine Bewertungen
- Basal ModificationDokument21 SeitenBasal ModificationPatrick Rómulo CabilingNoch keine Bewertungen
- Week 4 Case StudyDokument6 SeitenWeek 4 Case StudyElizabeth GithinjiNoch keine Bewertungen
- Skin Barrier Function - Morphological Basis and Regulatory MechanismsDokument10 SeitenSkin Barrier Function - Morphological Basis and Regulatory MechanismsJupiter CarlNoch keine Bewertungen
- Granulomatous Inflammations of Soft Tissue Include Tuberculosis, Atypical Mycobacteriosis, ActinomycosisDokument5 SeitenGranulomatous Inflammations of Soft Tissue Include Tuberculosis, Atypical Mycobacteriosis, ActinomycosisMișulică CerlatNoch keine Bewertungen
- Cell JunctionsDokument56 SeitenCell JunctionsDrGagandeep Singh100% (3)
- Anatomy, Skin (Integument), Epidermis - StatPearls - NCBI BookshelfDokument8 SeitenAnatomy, Skin (Integument), Epidermis - StatPearls - NCBI Bookshelfpka25Noch keine Bewertungen
- Skin Basic Structure and FunctionDokument11 SeitenSkin Basic Structure and FunctionPanggih Sekar Palupi IINoch keine Bewertungen
- Desmosoma y Hemodesmosoma PDFDokument11 SeitenDesmosoma y Hemodesmosoma PDFAna LabeNoch keine Bewertungen
- Ackerman - Histologic Diagnosis of Inflammatoy Skin Diseases - Third EditionDokument2.066 SeitenAckerman - Histologic Diagnosis of Inflammatoy Skin Diseases - Third EditionSamanthaNoch keine Bewertungen
- Harrys CosmetologyDokument80 SeitenHarrys CosmetologyNidhi100% (3)
- 1.integumentary System - HistologyDokument34 Seiten1.integumentary System - Histologyqty9jgkpnzNoch keine Bewertungen
- Meth Elab Topic N1Dokument23 SeitenMeth Elab Topic N1Group M1869Noch keine Bewertungen
- Skin Structure and FunctionDokument4 SeitenSkin Structure and FunctionPutri ClaraNoch keine Bewertungen
- The Role of Tight Junctions in Skin Barrier Function Anddermal AbsorptionDokument14 SeitenThe Role of Tight Junctions in Skin Barrier Function Anddermal AbsorptionPaolaNoch keine Bewertungen
- Cell JunctionsDokument40 SeitenCell JunctionsAhmad MursyidNoch keine Bewertungen
- Cell-Cell Connectivity: Desmosomes and Disease: Invited ReviewDokument14 SeitenCell-Cell Connectivity: Desmosomes and Disease: Invited ReviewMARIA ANGGIE CANTIKA DEWANINoch keine Bewertungen
- Cell JunctionsDokument29 SeitenCell JunctionsMira Cindy IINoch keine Bewertungen
- Dermatology Questions and AnsDokument147 SeitenDermatology Questions and AnsEl FaroukNoch keine Bewertungen
- Dermis Is A Specialized EpidermisDokument4 SeitenDermis Is A Specialized EpidermisfaisNoch keine Bewertungen
- Unit 1: Biochemistry 1.1 Plasma MembraneDokument12 SeitenUnit 1: Biochemistry 1.1 Plasma MembraneThrupthiNoch keine Bewertungen
- Bab 7. Struktur KulitDokument56 SeitenBab 7. Struktur KulityumizoneNoch keine Bewertungen
- Cell Adhesion Molecules: Mrs. Ofelia Solano SaludarDokument15 SeitenCell Adhesion Molecules: Mrs. Ofelia Solano SaludarmskikiNoch keine Bewertungen
- Cell Junctions: Mrs. Ofelia Solano SaludarDokument19 SeitenCell Junctions: Mrs. Ofelia Solano SaludarmskikiNoch keine Bewertungen
- The SkinDokument10 SeitenThe SkinTeriany WidjayaNoch keine Bewertungen
- JDD Atopic Dermatitis and The Role For A Ceramide Dominant Physiologic Lipid Based Barrier Repair EmulsionDokument5 SeitenJDD Atopic Dermatitis and The Role For A Ceramide Dominant Physiologic Lipid Based Barrier Repair EmulsionFajrin Dwi SyaputraNoch keine Bewertungen
- En Con Celladhesion 20120407Dokument17 SeitenEn Con Celladhesion 20120407CLPHtheoryNoch keine Bewertungen
- The Cis-Acting Signals That Target Proteins To Exosomes and MicrovesiclesDokument6 SeitenThe Cis-Acting Signals That Target Proteins To Exosomes and MicrovesiclesGlauce L TrevisanNoch keine Bewertungen
- Vertebrate Integument and DerivativesDokument16 SeitenVertebrate Integument and DerivativesNarasimha MurthyNoch keine Bewertungen
- Integumentary System: Stratum BasaleDokument21 SeitenIntegumentary System: Stratum BasalerinaNoch keine Bewertungen
- Lecture 27 - Histology of The Integumentary System 2Dokument89 SeitenLecture 27 - Histology of The Integumentary System 2spitzmark2030Noch keine Bewertungen
- 3 Cytology Membran JunctionDokument48 Seiten3 Cytology Membran JunctionSamNoch keine Bewertungen
- Skin Structure and Function: Figure 1.1 Cross-Section of The SkinDokument11 SeitenSkin Structure and Function: Figure 1.1 Cross-Section of The SkinfunmugNoch keine Bewertungen
- Integumentary SystemDokument24 SeitenIntegumentary SystemWadabiNoch keine Bewertungen
- Structure of Skin SeminarDokument65 SeitenStructure of Skin SeminarVikas PurushottamNoch keine Bewertungen
- Human Anatomy 5th Edition Saladin Solutions Manual 1Dokument36 SeitenHuman Anatomy 5th Edition Saladin Solutions Manual 1donnawugnwsjrzcxt100% (22)
- Human Anatomy 5Th Edition Saladin Solutions Manual Full Chapter PDFDokument28 SeitenHuman Anatomy 5Th Edition Saladin Solutions Manual Full Chapter PDFdora.ivy892100% (17)
- Human Anatomy 5th Edition Saladin Solutions Manual 1Dokument7 SeitenHuman Anatomy 5th Edition Saladin Solutions Manual 1katherine100% (41)
- Artikel 2 Regulation of Skin PigmentationDokument5 SeitenArtikel 2 Regulation of Skin PigmentationNandhy AlpNoch keine Bewertungen
- Oral Mucous MembraneDokument71 SeitenOral Mucous MembraneHarleen GrewalNoch keine Bewertungen
- Library DissertationDokument201 SeitenLibrary Dissertationhaneefmdf100% (2)
- Keratinization of The Oral EpitheliumDokument8 SeitenKeratinization of The Oral EpitheliumZachary DuongNoch keine Bewertungen
- 3 B Cytology-Membrane StructuresDokument19 Seiten3 B Cytology-Membrane Structuresm.mansoor3377Noch keine Bewertungen
- Cancer MecanismosDokument13 SeitenCancer MecanismosDianitaGaravitoVargasNoch keine Bewertungen
- Skin Structure and Function: Applied Dermatotoxicology. DOI: © 2014 Elsevier Inc. All Rights ReservedDokument10 SeitenSkin Structure and Function: Applied Dermatotoxicology. DOI: © 2014 Elsevier Inc. All Rights ReservedArsene AngelaNoch keine Bewertungen
- Write An Eassy On The Different Types of Cell Junctions That Occur in Plant and Animal TissuesDokument8 SeitenWrite An Eassy On The Different Types of Cell Junctions That Occur in Plant and Animal TissuestanatswaNoch keine Bewertungen
- Integrasi Sel Dalam Jaringan (Kuliah) - 2 - 2Dokument125 SeitenIntegrasi Sel Dalam Jaringan (Kuliah) - 2 - 2komang nickoNoch keine Bewertungen
- Skin Anatomy and Flap Physiology: P. S. MDDokument15 SeitenSkin Anatomy and Flap Physiology: P. S. MDAncaVerdesNoch keine Bewertungen
- Cytokines and The Skin Barrier: Molecular SciencesDokument26 SeitenCytokines and The Skin Barrier: Molecular SciencesFelp ScholzNoch keine Bewertungen
- Epidermal Sphingolipids: Metabolism, Function, and Roles in Skin DisordersDokument11 SeitenEpidermal Sphingolipids: Metabolism, Function, and Roles in Skin DisorderspangeiaaNoch keine Bewertungen
- 1 s2.0 S0022202X15447736 MainDokument7 Seiten1 s2.0 S0022202X15447736 MainKhan FatemaNoch keine Bewertungen
- Anatomy and PhysiologyDokument25 SeitenAnatomy and PhysiologyNaidin Catherine De Guzman-AlcalaNoch keine Bewertungen
- Christersson2022 Article ScreeningForBiomarkersAssociatDokument11 SeitenChristersson2022 Article ScreeningForBiomarkersAssociatS FznsNoch keine Bewertungen
- Clinical Relevance of Elastin in The Structure and Function of SkinDokument8 SeitenClinical Relevance of Elastin in The Structure and Function of SkinS FznsNoch keine Bewertungen
- Infant Skin Barrier, Structure, and Enzymatic Activity Differ From Those of Adult in An East Asian CohortDokument8 SeitenInfant Skin Barrier, Structure, and Enzymatic Activity Differ From Those of Adult in An East Asian CohortS FznsNoch keine Bewertungen
- Prurigo Simplex or Itchy Red BumpsDokument7 SeitenPrurigo Simplex or Itchy Red BumpsS FznsNoch keine Bewertungen
- Clinical and Radiological Outcomes of Infantile Hemangioma Treated With Oral Propranolol: A Long-Term Follow-Up StudyDokument7 SeitenClinical and Radiological Outcomes of Infantile Hemangioma Treated With Oral Propranolol: A Long-Term Follow-Up StudyS FznsNoch keine Bewertungen
- Onychomycosis An Updated ReviewDokument14 SeitenOnychomycosis An Updated ReviewS FznsNoch keine Bewertungen
- 38th APAO Congress E Poster Presentation GuidelinesDokument3 Seiten38th APAO Congress E Poster Presentation GuidelinesS FznsNoch keine Bewertungen
- BAHAN Chapter 24Dokument33 SeitenBAHAN Chapter 24S FznsNoch keine Bewertungen
- Minity General Menu - 2022-1Dokument10 SeitenMinity General Menu - 2022-1S FznsNoch keine Bewertungen
- Methodology Checklist 1: Systematic Reviews and Meta-AnalysesDokument2 SeitenMethodology Checklist 1: Systematic Reviews and Meta-AnalysesS FznsNoch keine Bewertungen
- Sisa Rangkum SalmonellaDokument2 SeitenSisa Rangkum SalmonellaS FznsNoch keine Bewertungen
- JamaDokument4 SeitenJamaS FznsNoch keine Bewertungen
- Medicina: Oral and Maxillo-Facial Manifestations of Systemic Diseases: An OverviewDokument22 SeitenMedicina: Oral and Maxillo-Facial Manifestations of Systemic Diseases: An OverviewS FznsNoch keine Bewertungen
- BAHAN Fitzpatrick 13 - 2Dokument27 SeitenBAHAN Fitzpatrick 13 - 2S FznsNoch keine Bewertungen
- Liver AbsesDokument8 SeitenLiver AbsesS FznsNoch keine Bewertungen
- The Concept of Soluble Mediators: Tumor Necrosis FactorDokument4 SeitenThe Concept of Soluble Mediators: Tumor Necrosis FactorS FznsNoch keine Bewertungen
- Chapter 6:: Skin Glands: Sebaceous, Eccrine, and Apocrine GlandsDokument32 SeitenChapter 6:: Skin Glands: Sebaceous, Eccrine, and Apocrine GlandsS FznsNoch keine Bewertungen
- Chapter 13:: Basic Principles of Immunologic Diseases in Skin (Pathophysiology of Immunologic/Inflammatory Skin Diseases)Dokument7 SeitenChapter 13:: Basic Principles of Immunologic Diseases in Skin (Pathophysiology of Immunologic/Inflammatory Skin Diseases)S FznsNoch keine Bewertungen
- BAHAN Infanticide ConceptDokument5 SeitenBAHAN Infanticide ConceptS FznsNoch keine Bewertungen
- Overactive Bladder Syndrome: Management and Treatment OptionsDokument7 SeitenOveractive Bladder Syndrome: Management and Treatment OptionsS FznsNoch keine Bewertungen
- Bab I-V Jurding Infanticide EditedDokument62 SeitenBab I-V Jurding Infanticide EditedS FznsNoch keine Bewertungen
- Preterm Preeclampsia and Timing of DeliveryDokument3 SeitenPreterm Preeclampsia and Timing of DeliveryS FznsNoch keine Bewertungen
- Epifix BrochureDokument2 SeitenEpifix Brochureapi-235072265100% (2)
- CHAPTER 3. Tissue BiologyDokument26 SeitenCHAPTER 3. Tissue Biologywella wellaNoch keine Bewertungen
- Week 4 Tissue Levels of OrganizatioDokument49 SeitenWeek 4 Tissue Levels of OrganizatioDayledaniel SorvetoNoch keine Bewertungen
- Karps Cell and Molecular Biology Ninth 9Th Edition Gerald Karp Full ChapterDokument67 SeitenKarps Cell and Molecular Biology Ninth 9Th Edition Gerald Karp Full Chapterronald.dancy562100% (8)
- Corneal Epithelial Wound Healing: PerspectiveDokument8 SeitenCorneal Epithelial Wound Healing: PerspectiveHanaNoch keine Bewertungen
- Junctional Epithelium: Dr. Ishita Singhal Mds First YearDokument54 SeitenJunctional Epithelium: Dr. Ishita Singhal Mds First YearDR. ISHITA SINGHALNoch keine Bewertungen
- Pearls of Glaucoma ManagementDokument576 SeitenPearls of Glaucoma Managementrobeye100% (1)
- Epifix Amniofix InformationDokument11 SeitenEpifix Amniofix Informationapi-235072265100% (1)
- Basf Presentation Radiant SkinDokument39 SeitenBasf Presentation Radiant SkinJames WuNoch keine Bewertungen
- Quiz #1 Questions A&P 1Dokument14 SeitenQuiz #1 Questions A&P 1trick49Noch keine Bewertungen
- Animal TissuesDokument53 SeitenAnimal TissuesIpshita PathakNoch keine Bewertungen
- Histology: Lecture NotesDokument232 SeitenHistology: Lecture NotesAdrian PuscasNoch keine Bewertungen
- Oral Mucosal Drug Delivery and TherapyDokument289 SeitenOral Mucosal Drug Delivery and TherapyАлександр ВолошанNoch keine Bewertungen
- Histology Prelim NotesDokument7 SeitenHistology Prelim NotesKathleen Cobrador EribalNoch keine Bewertungen
- Epithelium IDokument33 SeitenEpithelium IUmerNoch keine Bewertungen
- Junctional Epithelium: Presented By: Dr. Monali B.Pimple 1 Year PG StudentDokument40 SeitenJunctional Epithelium: Presented By: Dr. Monali B.Pimple 1 Year PG Studentmonali pimple50% (2)
- The Cornea: Anatomy & PhysiologyDokument24 SeitenThe Cornea: Anatomy & PhysiologyLeo TedjaNoch keine Bewertungen
- Skin AnatomyDokument71 SeitenSkin AnatomyHardik DaveNoch keine Bewertungen
- Cancer Biology Assignment: SR - NO. Topic Roll No. NameDokument21 SeitenCancer Biology Assignment: SR - NO. Topic Roll No. NameSimran PhutelaNoch keine Bewertungen
- Anatomy of Cornea and Ocular Surface.4Dokument5 SeitenAnatomy of Cornea and Ocular Surface.4Cucută Alexandru-DanielNoch keine Bewertungen
- CAM101 Complete Course Notes Reduced SizeDokument174 SeitenCAM101 Complete Course Notes Reduced SizehaydenfarquharNoch keine Bewertungen
- Molecular & Cell Biology of GingivaDokument28 SeitenMolecular & Cell Biology of GingivaPoojan ThakoreNoch keine Bewertungen
- Epithelium I (The First of Four Tissues) : Reading Assignment: Junqueira & Carneiro, Chapt. 2&4 Gartner & Hiatt, Chapter 2Dokument57 SeitenEpithelium I (The First of Four Tissues) : Reading Assignment: Junqueira & Carneiro, Chapt. 2&4 Gartner & Hiatt, Chapter 2iramfarazNoch keine Bewertungen
- Histology Exam I OutlineDokument76 SeitenHistology Exam I OutlinePrinceNoch keine Bewertungen
- Simple EpitheliumDokument64 SeitenSimple EpitheliumRajesh KumarNoch keine Bewertungen
- Awdal College Department of Health: General HistologyDokument24 SeitenAwdal College Department of Health: General HistologyAyro Business CenterNoch keine Bewertungen
- Neonatal SkinDokument384 SeitenNeonatal SkinErnestas ViršilasNoch keine Bewertungen
- Part 7Dokument7 SeitenPart 7Anna Marie Ventayen MirandaNoch keine Bewertungen
- Palate Development. Ferguson.Dokument20 SeitenPalate Development. Ferguson.Fabian BarretoNoch keine Bewertungen
- Geltrex Culture Medium GuideDokument2 SeitenGeltrex Culture Medium GuideJohn ExtonNoch keine Bewertungen