Das könnte Ihnen auch gefallen
- Physiology Practical 2: Toad HeartDokument10 SeitenPhysiology Practical 2: Toad HeartAdams OdanjiNoch keine Bewertungen
- Lab Report 2 AutosavedDokument18 SeitenLab Report 2 Autosavedapi-249482502100% (2)
- Effects of Temperature, Stretch, and Various Drug Treatments On The Cardiac Muscle Activity of Rana PipiensDokument14 SeitenEffects of Temperature, Stretch, and Various Drug Treatments On The Cardiac Muscle Activity of Rana PipiensMereseini VakamacalaNoch keine Bewertungen
- Lab08 Frog HeartDokument4 SeitenLab08 Frog HeartAbie CaponponNoch keine Bewertungen
- Bio 122 FR 10 - Cardiac MuscleDokument11 SeitenBio 122 FR 10 - Cardiac MuscleLance CarandangNoch keine Bewertungen
- CardiovascularDokument16 SeitenCardiovascularMilena VargasNoch keine Bewertungen
- DocxDokument18 SeitenDocxmutindaNoch keine Bewertungen
- Cardiac Cycle Phases and Effects of Temperature, ChemicalsDokument6 SeitenCardiac Cycle Phases and Effects of Temperature, ChemicalsMatthew SANoch keine Bewertungen
- Properties of Cardiac MuscleDokument5 SeitenProperties of Cardiac Musclekiedd_04100% (4)
- Lab 5 ReportDokument19 SeitenLab 5 Reportapi-248983324Noch keine Bewertungen
- The Physiology of A Frog Heart With The Effects of Temperature, Neurotransmitters, and Electrical IsolationDokument17 SeitenThe Physiology of A Frog Heart With The Effects of Temperature, Neurotransmitters, and Electrical IsolationAbir Hasan ShuvoNoch keine Bewertungen
- Effects of Temperature, Stretch, and Various Drug Treatments On The Cardiac Muscle Activity of Rana PipiensDokument15 SeitenEffects of Temperature, Stretch, and Various Drug Treatments On The Cardiac Muscle Activity of Rana PipiensAbir Hasan ShuvoNoch keine Bewertungen
- Cardiac cycle: Mechanical events and hemodynamic changesDokument20 SeitenCardiac cycle: Mechanical events and hemodynamic changesKelly Yeow100% (3)
- CVS ChartsDokument67 SeitenCVS Chartssacin99999Noch keine Bewertungen
- Lab Report 5 - Yixi LiuDokument22 SeitenLab Report 5 - Yixi Liuapi-308855010100% (1)
- Circulation and Cardiovascular SystemDokument12 SeitenCirculation and Cardiovascular SystemSatria MandalaNoch keine Bewertungen
- Physiocardiacoutput 171108041156Dokument20 SeitenPhysiocardiacoutput 171108041156Akila AkinsNoch keine Bewertungen
- CV SystemDokument31 SeitenCV SystemaletheacolleenpNoch keine Bewertungen
- Cardiovascular System Physiology, Lecture 2 (Cardiodynamics)Dokument15 SeitenCardiovascular System Physiology, Lecture 2 (Cardiodynamics)Sherwan R Shal100% (5)
- Ventricular Contraction Larger Waves OscilloscopeDokument5 SeitenVentricular Contraction Larger Waves Oscilloscopeskyhigh299100% (2)
- Student ProtocolDokument14 SeitenStudent ProtocolLaura Camila Bermeo PeraltaNoch keine Bewertungen
- Cardiovascular Physiology: The Autonomic Nervous SystemDokument8 SeitenCardiovascular Physiology: The Autonomic Nervous SystemRidha Surya NugrahaNoch keine Bewertungen
- Physiology of The HeartDokument34 SeitenPhysiology of The Heartalyssa_marie_keNoch keine Bewertungen
- Physiology - Cardiovascular SystemDokument16 SeitenPhysiology - Cardiovascular SystemHey GoogleNoch keine Bewertungen
- Digital Heart Beat Counter: Project Report ONDokument76 SeitenDigital Heart Beat Counter: Project Report ONlkosumdecNoch keine Bewertungen
- PhysioEx Exercise 5Dokument21 SeitenPhysioEx Exercise 5Sinisa Ristic100% (1)
- Monash Physiology Simulation Cardiac Muscle LengthDokument25 SeitenMonash Physiology Simulation Cardiac Muscle Lengthluis11256Noch keine Bewertungen
- ECG Pulse Physiology Lab ExperimentsDokument3 SeitenECG Pulse Physiology Lab Experimentsaileen agustin100% (6)
- Control of Stroke VolumeDokument2 SeitenControl of Stroke VolumeNanda RahmatNoch keine Bewertungen
- Lab #4: Physiology of The in Situ Amphibian Heart: BackgroundDokument6 SeitenLab #4: Physiology of The in Situ Amphibian Heart: BackgroundAbir Hasan ShuvoNoch keine Bewertungen
- BP503T Unit 1-3Dokument432 SeitenBP503T Unit 1-3Solomon GyampohNoch keine Bewertungen
- Frank-Starling LawDokument6 SeitenFrank-Starling LawGauri RastogiNoch keine Bewertungen
- Heart Rate Blood PressureDokument128 SeitenHeart Rate Blood PressureShubhra ShettyNoch keine Bewertungen
- Study For MCB CardioDokument12 SeitenStudy For MCB Cardiolavanya.aNoch keine Bewertungen
- ECG & Heart Sounds: Student HandoutDokument7 SeitenECG & Heart Sounds: Student HandoutROHITNoch keine Bewertungen
- Student ProtocolDokument14 SeitenStudent ProtocolCristian AmadoNoch keine Bewertungen
- Evaluation of Pumping Function of Heart: Stroke Volume Cardiac Output Ejection Fraction Cardiac IndexDokument53 SeitenEvaluation of Pumping Function of Heart: Stroke Volume Cardiac Output Ejection Fraction Cardiac Indexapi-19916399Noch keine Bewertungen
- Frog Heart - Tharp QuestionsDokument2 SeitenFrog Heart - Tharp QuestionscrabcroutonNoch keine Bewertungen
- Lecture - 3 Properties of Cardiac MuscleDokument35 SeitenLecture - 3 Properties of Cardiac MuscleMRM7MDNoch keine Bewertungen
- Ecg Heart Sounds Laboratory HandoutDokument7 SeitenEcg Heart Sounds Laboratory HandoutShashank SahuNoch keine Bewertungen
- Frank-Starling LawDokument5 SeitenFrank-Starling LawNTA UGC-NETNoch keine Bewertungen
- 51456-CVS-23086215D-Cheung Chi Ming.doc - 複本Dokument6 Seiten51456-CVS-23086215D-Cheung Chi Ming.doc - 複本9rw9yrh6qyNoch keine Bewertungen
- ECG & Heart Sounds LabDokument7 SeitenECG & Heart Sounds LabPatrick Joshua PascualNoch keine Bewertungen
- Physiology: Zhao Chun-ling (赵春玲) Email: Phone:18683073667Dokument62 SeitenPhysiology: Zhao Chun-ling (赵春玲) Email: Phone:18683073667Akshay ChaudharyNoch keine Bewertungen
- Cardiac Output HandoutDokument10 SeitenCardiac Output Handoutsac50900Noch keine Bewertungen
- Cardiac Cycle Option HDokument8 SeitenCardiac Cycle Option HstorykuNoch keine Bewertungen
- Measure tidal volume and rate of breathingDokument8 SeitenMeasure tidal volume and rate of breathingRuwan BandaraNoch keine Bewertungen
- The Normal Physiology of The Heart: Vikkineshwaran Siva SubramaniamDokument4 SeitenThe Normal Physiology of The Heart: Vikkineshwaran Siva SubramaniamSivamala MalaNoch keine Bewertungen
- 51456-CVS-23086215D-Cheung-Chi-Ming.docDokument6 Seiten51456-CVS-23086215D-Cheung-Chi-Ming.doc9rw9yrh6qyNoch keine Bewertungen
- 25circulation Part 3Dokument15 Seiten25circulation Part 3Jaydave PatelNoch keine Bewertungen
- Cardiovascular System Chapter SummaryDokument4 SeitenCardiovascular System Chapter Summarycuksam27Noch keine Bewertungen
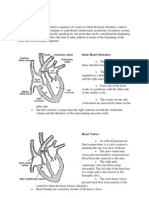
- Cardiac Cycle.: Basic Heart StructureDokument9 SeitenCardiac Cycle.: Basic Heart StructureMohammad AbdullahNoch keine Bewertungen
- EECP IabpDokument16 SeitenEECP IabpMythily VedhagiriNoch keine Bewertungen
- Cardiovascular Physiology Practice QuestionsDokument4 SeitenCardiovascular Physiology Practice QuestionsHayley Welsh100% (1)
- The 12-Lead Electrocardiogram for Nurses and Allied ProfessionalsVon EverandThe 12-Lead Electrocardiogram for Nurses and Allied ProfessionalsNoch keine Bewertungen
- Preload and AfterloadDokument28 SeitenPreload and Afterloadapi-19916399100% (1)
- Cardiac Muscle Refractory PeriodDokument7 SeitenCardiac Muscle Refractory PeriodEngku FaisalNoch keine Bewertungen
- Outline The Transmission of The Electrical Impulse Through The HeartDokument3 SeitenOutline The Transmission of The Electrical Impulse Through The HeartHannah JohnstonNoch keine Bewertungen
- Central Venous Pressure: Its Clinical Use and Role in Cardiovascular DynamicsVon EverandCentral Venous Pressure: Its Clinical Use and Role in Cardiovascular DynamicsNoch keine Bewertungen
- High Blood Pressure: Safe alternatives without drugsVon EverandHigh Blood Pressure: Safe alternatives without drugsBewertung: 5 von 5 Sternen5/5 (2)
- Dispersal Report FinalDokument16 SeitenDispersal Report FinalAlex HebertNoch keine Bewertungen
- Lecture 1Dokument97 SeitenLecture 1Alex HebertNoch keine Bewertungen
- Control of Microbial GrowthDokument4 SeitenControl of Microbial GrowthAlex HebertNoch keine Bewertungen
- Hypothesis Cardiac LabDokument2 SeitenHypothesis Cardiac LabAlex HebertNoch keine Bewertungen
- Lecture 1 IntroductionDokument44 SeitenLecture 1 IntroductionAlex HebertNoch keine Bewertungen
- How temperature and pressure affect chemical equilibriumDokument1 SeiteHow temperature and pressure affect chemical equilibriumAlex HebertNoch keine Bewertungen
- MicroDokument44 SeitenMicroAlex HebertNoch keine Bewertungen
- PacmakerDokument37 SeitenPacmakerSalman HabeebNoch keine Bewertungen
- Management of Cardiogenic ShockDokument15 SeitenManagement of Cardiogenic ShockcastillojessNoch keine Bewertungen
- Quiz 8 To 46Dokument7 SeitenQuiz 8 To 46JhayneNoch keine Bewertungen
- Hypertension CaseDokument3 SeitenHypertension CaseArnold Christian QuilonNoch keine Bewertungen
- Indications: PurposeDokument13 SeitenIndications: PurposeIRA ODETTE GATUSNoch keine Bewertungen
- The Pharmacology of Adrenergic Receptors PDFDokument15 SeitenThe Pharmacology of Adrenergic Receptors PDFKathryn MitchellNoch keine Bewertungen
- Hypoxic SpellsDokument3 SeitenHypoxic SpellsMemo LoezaNoch keine Bewertungen
- ECG PG 46 ST Elevation Smiley Early Repol (7 17.1 2011) LOCKDokument0 SeitenECG PG 46 ST Elevation Smiley Early Repol (7 17.1 2011) LOCKJD DaguioNoch keine Bewertungen
- The Artery of Adamkiewicz Vascular Anatomy Clinical Significance and Surgical Considerations 9VoDDokument5 SeitenThe Artery of Adamkiewicz Vascular Anatomy Clinical Significance and Surgical Considerations 9VoDNitishNoch keine Bewertungen
- MagdiDokument4 SeitenMagdiapi-534579275Noch keine Bewertungen
- Vitality Fitness Assessment Form: 1. DetailsDokument2 SeitenVitality Fitness Assessment Form: 1. DetailsmusturNoch keine Bewertungen
- Cardiac Murmurs in Children: FactfileDokument2 SeitenCardiac Murmurs in Children: FactfileResya I. NoerNoch keine Bewertungen
- Untitled19 PDFDokument19 SeitenUntitled19 PDFElizabeth LeonNoch keine Bewertungen
- Vasopressor Use in Adult Patients: Nadia Ferguson-Myrthil, PharmdDokument6 SeitenVasopressor Use in Adult Patients: Nadia Ferguson-Myrthil, PharmdWILLIAM RICARDO CASTAÑEDA VARGASNoch keine Bewertungen
- CCD 25132Dokument3 SeitenCCD 25132bharattaneja017023Noch keine Bewertungen
- Health Teachings: (Select All That Apply)Dokument6 SeitenHealth Teachings: (Select All That Apply)bon clayNoch keine Bewertungen
- Heart FailureDokument118 SeitenHeart FailureAli Murtaza AbbasNoch keine Bewertungen
- Myocardial InfarctionDokument3 SeitenMyocardial Infarctionaja_delNoch keine Bewertungen
- Analisis Cost of Illness Penyakit Hipertensi Di Rumah Sakit Umum Daerah Praya, NTBDokument8 SeitenAnalisis Cost of Illness Penyakit Hipertensi Di Rumah Sakit Umum Daerah Praya, NTBabdulNoch keine Bewertungen
- Transient Ischemic Attack Precipitating Factors Predisposing FactorsDokument6 SeitenTransient Ischemic Attack Precipitating Factors Predisposing FactorsYosef OxinioNoch keine Bewertungen
- 10 Peri Arrest ArrhythmiasDokument39 Seiten10 Peri Arrest ArrhythmiasAnaid FourevaNoch keine Bewertungen
- (Junjie Xiao (Eds.) ) Exercise For Cardiovascular D (B-Ok - CC)Dokument380 Seiten(Junjie Xiao (Eds.) ) Exercise For Cardiovascular D (B-Ok - CC)Nacido para BendcirNoch keine Bewertungen
- Heart Failure Clinical Presentation - History, Physical Examination, Predominant Right-Sided Heart Failure PDFDokument11 SeitenHeart Failure Clinical Presentation - History, Physical Examination, Predominant Right-Sided Heart Failure PDFislamiah 89Noch keine Bewertungen
- Effect of Caffeine and Nicotine On Blood Pressure.Dokument1 SeiteEffect of Caffeine and Nicotine On Blood Pressure.Mohammed Danish AliNoch keine Bewertungen
- Nurses ECG - 1Dokument72 SeitenNurses ECG - 1Aswini Kumar SNoch keine Bewertungen
- Philips HeartStart XLPlus Owners Manual PDFDokument234 SeitenPhilips HeartStart XLPlus Owners Manual PDFmogfacNoch keine Bewertungen
- Part1 - CIE IGCSE Biology 9 RN - SampleDokument2 SeitenPart1 - CIE IGCSE Biology 9 RN - SampleJun ZheNoch keine Bewertungen
- Swallow SyncopeDokument4 SeitenSwallow SyncopeKevinSheridanNoch keine Bewertungen
- Journal Reading: Imaging of Intracranial HemorrhageDokument27 SeitenJournal Reading: Imaging of Intracranial HemorrhagehasrilNoch keine Bewertungen
- Left Ventricle Wall MotionDokument4 SeitenLeft Ventricle Wall MotionSolenero SatelNoch keine Bewertungen