Das könnte Ihnen auch gefallen
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
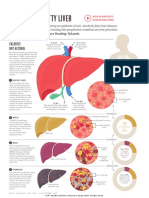
- Fatty LiverDokument2 SeitenFatty LiverKarla Edith Rodriguez Nava100% (2)
- Intro BotanyDokument192 SeitenIntro BotanyMelissa100% (1)
- Kalish GI ProtocolsDokument5 SeitenKalish GI Protocolsgoosenl100% (1)
- 3rd Term SS1Dokument31 Seiten3rd Term SS1ofurumchinyere9Noch keine Bewertungen
- Life Science Previous Paper 2017 DecDokument38 SeitenLife Science Previous Paper 2017 DecShilpi PriyaNoch keine Bewertungen
- Epigenetic and Cancer Therapy, 2019Dokument12 SeitenEpigenetic and Cancer Therapy, 2019AbsjeyNoch keine Bewertungen
- Relevanttopicalskincare Productsforprevention AndtreatmentofagingskinDokument6 SeitenRelevanttopicalskincare Productsforprevention AndtreatmentofagingskinenviNoch keine Bewertungen
- Fine Test ELISA KitsDokument376 SeitenFine Test ELISA KitsDavid1002Noch keine Bewertungen
- Enzymatic Cleavage and The Creation of Heteroduplex DNA: Holliday ModelDokument10 SeitenEnzymatic Cleavage and The Creation of Heteroduplex DNA: Holliday ModelAshwini PillaiNoch keine Bewertungen
- NSW Biology Year 11 Question and Answer Booklet 2022Dokument24 SeitenNSW Biology Year 11 Question and Answer Booklet 2022annaNoch keine Bewertungen
- Sathyanarayan Biochemistry 4epdfDokument35 SeitenSathyanarayan Biochemistry 4epdfAbubakar SuleimanNoch keine Bewertungen
- Session #43 SAS - AnaPhy (Lab)Dokument4 SeitenSession #43 SAS - AnaPhy (Lab)G INoch keine Bewertungen
- Blood: Blood Transports Everything That Must Be Carried From One Place To Another, Such AsDokument71 SeitenBlood: Blood Transports Everything That Must Be Carried From One Place To Another, Such AsAudrie Allyson GabalesNoch keine Bewertungen
- Microbiology An Introduction 11Th Edition Tortora Test Bank Full Chapter PDFDokument42 SeitenMicrobiology An Introduction 11Th Edition Tortora Test Bank Full Chapter PDFemilymorrowjmtafrqcdn100% (9)
- FAQs: Dr. Sarojini SahooDokument4 SeitenFAQs: Dr. Sarojini SahooSarojini SahooNoch keine Bewertungen
- Daily Lesson Log Senior High School: I. ObjectivesDokument3 SeitenDaily Lesson Log Senior High School: I. ObjectivesAl-John EspejoNoch keine Bewertungen
- Biotech Project BC0170027Dokument10 SeitenBiotech Project BC0170027Shretha lakshmi rNoch keine Bewertungen
- 823-Article Text-3390-2-10-20190624Dokument12 Seiten823-Article Text-3390-2-10-20190624umar yusufNoch keine Bewertungen
- Pre Biology POSN - Examination PLUSDokument108 SeitenPre Biology POSN - Examination PLUSTetee TetatNoch keine Bewertungen
- Environment VS SocietyDokument9 SeitenEnvironment VS SocietyPriyanka PalNoch keine Bewertungen
- Clostridium Botulinum: Molecular Medicines For Neutralization of NeurotoxinDokument13 SeitenClostridium Botulinum: Molecular Medicines For Neutralization of NeurotoxinArshia NazirNoch keine Bewertungen
- Pathogenesis of Type 1 Diabetes MellitusDokument12 SeitenPathogenesis of Type 1 Diabetes MellitusJIEHASMARTNoch keine Bewertungen
- Microbial Carcinogenesis: Dr. Kevin Manuel Assistant Professor Department of Pathology 19.09.18Dokument37 SeitenMicrobial Carcinogenesis: Dr. Kevin Manuel Assistant Professor Department of Pathology 19.09.18Chama TlauNoch keine Bewertungen
- Module 14Dokument9 SeitenModule 14mirai desuNoch keine Bewertungen
- Endocrine System: Organs and Their FunctionDokument61 SeitenEndocrine System: Organs and Their FunctionSherifaNoch keine Bewertungen
- Principles of Ecology Study Guide Test 3 Temple UnivDokument8 SeitenPrinciples of Ecology Study Guide Test 3 Temple UnivChris VelasquezNoch keine Bewertungen
- NanoCellect Biomedical Begins Distribution To EuropeDokument2 SeitenNanoCellect Biomedical Begins Distribution To EuropePR.comNoch keine Bewertungen
- Penawaran Harga PCR KitDokument2 SeitenPenawaran Harga PCR KithafizauliaNoch keine Bewertungen
- Cyclin A Protein Specified by Maternal MRNA in Sea Urchin Eggs That Is Destroyed at Each Cleavage DivisionDokument8 SeitenCyclin A Protein Specified by Maternal MRNA in Sea Urchin Eggs That Is Destroyed at Each Cleavage Divisionjavi0% (1)
- Hem Lab Manual Diff F16Dokument10 SeitenHem Lab Manual Diff F16LAB.RSWB0% (1)