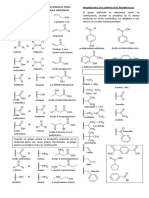
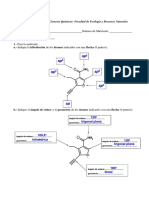
Das könnte Ihnen auch gefallen
- Comparación Multiple y Prueba de OrdenamientoDokument2 SeitenComparación Multiple y Prueba de OrdenamientoYCristina AcostaNoch keine Bewertungen
- Volumenes Parciales MolaresDokument4 SeitenVolumenes Parciales MolaresYCristina AcostaNoch keine Bewertungen
- Tecnicas de Evaluación de Gustos PrimariosDokument8 SeitenTecnicas de Evaluación de Gustos PrimariosYCristina AcostaNoch keine Bewertungen
- Higiene FosfatoDokument6 SeitenHigiene FosfatoYCristina AcostaNoch keine Bewertungen
- Caracterización de Los Postulados Sobre La Teoria de AseguramientoDokument4 SeitenCaracterización de Los Postulados Sobre La Teoria de AseguramientoYCristina AcostaNoch keine Bewertungen
- Excreción de Nitrogeno en Las AvesDokument1 SeiteExcreción de Nitrogeno en Las AvesYCristina AcostaNoch keine Bewertungen
- Complejos ProductivosDokument2 SeitenComplejos ProductivosYCristina AcostaNoch keine Bewertungen
- Minijardin IntroducciónDokument2 SeitenMinijardin IntroducciónYCristina AcostaNoch keine Bewertungen
- Cava o Camara de RefrigeraciónDokument10 SeitenCava o Camara de RefrigeraciónYCristina AcostaNoch keine Bewertungen
- Aplicación de La Ecuación de Bernoulli y El Principio de TorricelliDokument11 SeitenAplicación de La Ecuación de Bernoulli y El Principio de TorricelliYCristina AcostaNoch keine Bewertungen
- Mini JardinesDokument6 SeitenMini JardinesYCristina AcostaNoch keine Bewertungen
- Prueba Triangular y Comparación PareadaDokument3 SeitenPrueba Triangular y Comparación PareadaYCristina AcostaNoch keine Bewertungen
- Entorno GerencialDokument3 SeitenEntorno GerencialYCristina AcostaNoch keine Bewertungen
- Como Se Clasifica El Café Como BebidaDokument4 SeitenComo Se Clasifica El Café Como BebidaYCristina AcostaNoch keine Bewertungen
- Entorno GerencialDokument3 SeitenEntorno GerencialYCristina AcostaNoch keine Bewertungen
- Obtención de FurfuralDokument2 SeitenObtención de FurfuralclonqfbNoch keine Bewertungen
- Halogenacion - Ramirez Gomez Juan PabloDokument2 SeitenHalogenacion - Ramirez Gomez Juan PabloJuan RamirezNoch keine Bewertungen
- El CarbonoDokument3 SeitenEl CarbonoNinfa BetancourtNoch keine Bewertungen
- MacromoleculasDokument44 SeitenMacromoleculasMiguel Angel Rodas Herrera89% (9)
- Lipidos 2Dokument11 SeitenLipidos 2REYES MONTENEGRO MIRANDANoch keine Bewertungen
- Tema7 BetaoxidacionDokument24 SeitenTema7 BetaoxidacionLucasNoch keine Bewertungen
- Practica 7 Reacciones HidrocarburosDokument11 SeitenPractica 7 Reacciones HidrocarburosAldo Aguilera100% (2)
- Semana #3. ALCOHOLES Y FENOLESDokument57 SeitenSemana #3. ALCOHOLES Y FENOLESAceite De ChocolateNoch keine Bewertungen
- Síntesis y Propiedades de PolímerosDokument3 SeitenSíntesis y Propiedades de PolímerosJOSE ANTONIO HERNANDEZ VERANoch keine Bewertungen
- Informe de OleofinasDokument58 SeitenInforme de OleofinasVeronica Richardson100% (1)
- Taller CH y LípidosDokument3 SeitenTaller CH y LípidosCristian Vergara100% (1)
- Plan Global Labo Orga 2Dokument28 SeitenPlan Global Labo Orga 2willan paredez mamaniNoch keine Bewertungen
- Cuestionario Practica 1Dokument4 SeitenCuestionario Practica 1BryanSantiagoSánchezNoch keine Bewertungen
- Nitrocompuestos Clase 34Dokument6 SeitenNitrocompuestos Clase 34halina pradaNoch keine Bewertungen
- AMINOÁCIDOSDokument13 SeitenAMINOÁCIDOSjosseline adriana rivera revollo100% (1)
- Prioridades de Grupos Funcionales Prioridades y Grupos para La Nomenclatura en Química Orgánica PDFDokument1 SeitePrioridades de Grupos Funcionales Prioridades y Grupos para La Nomenclatura en Química Orgánica PDFCamila Jimenez Bejarano0% (1)
- QM-2421 Quimica Orgánica IDokument2 SeitenQM-2421 Quimica Orgánica IGuery SaenzNoch keine Bewertungen
- Poster (Alcanos Ramificados)Dokument5 SeitenPoster (Alcanos Ramificados)Jimena MoralesNoch keine Bewertungen
- Semana 4 Lipidos y Acidos NucleicosDokument15 SeitenSemana 4 Lipidos y Acidos NucleicosLorena Canchaya VegaNoch keine Bewertungen
- Acetileno Informe FinalDokument10 SeitenAcetileno Informe Finalfabio boteroNoch keine Bewertungen
- Química OrgánicaDokument10 SeitenQuímica OrgánicaJhonatan Micha SilvaNoch keine Bewertungen
- Practica 02 - SaponificacionDokument13 SeitenPractica 02 - SaponificacionRutth Apolinario Rico100% (1)
- Pautas P Química Orgánica IDokument53 SeitenPautas P Química Orgánica IIgnacio ZamudioNoch keine Bewertungen
- Taller Nomenclatura en Química OrgánicaDokument10 SeitenTaller Nomenclatura en Química OrgánicaEmily IturraldeNoch keine Bewertungen
- 12 Compuestos OrganicosDokument122 Seiten12 Compuestos OrganicosNicolas Patricio SitjaNoch keine Bewertungen
- Práctica 2Dokument10 SeitenPráctica 2José Luis ValenciaNoch keine Bewertungen
- Haluros OrgánicosDokument4 SeitenHaluros OrgánicosRossi Achata Merlin100% (1)
- 6to QUIMICA 3er TRIM PDFDokument14 Seiten6to QUIMICA 3er TRIM PDFAlex CanduryNoch keine Bewertungen
- Bioquimica 40 Biomuleculas Milany GomezDokument7 SeitenBioquimica 40 Biomuleculas Milany GomezMila Gomez BadilloNoch keine Bewertungen
- Laboratorio 4 de Quimica Organica 2Dokument18 SeitenLaboratorio 4 de Quimica Organica 2api-277522779Noch keine Bewertungen