Das könnte Ihnen auch gefallen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- HHS Public Access: Cognitive Decline and Dementia in Diabetes: Mechanisms and Clinical ImplicationsDokument27 SeitenHHS Public Access: Cognitive Decline and Dementia in Diabetes: Mechanisms and Clinical ImplicationsSaddam FuadNoch keine Bewertungen
- Complementary Therapies in MedicineDokument4 SeitenComplementary Therapies in MedicineSaddam FuadNoch keine Bewertungen
- Hypertension and The Risk of Dementia: Cristina SierraDokument7 SeitenHypertension and The Risk of Dementia: Cristina SierraSaddam FuadNoch keine Bewertungen
- HHS Public Access: Cognitive Decline and Dementia in Diabetes: Mechanisms and Clinical ImplicationsDokument27 SeitenHHS Public Access: Cognitive Decline and Dementia in Diabetes: Mechanisms and Clinical ImplicationsSaddam FuadNoch keine Bewertungen
- Zhang 2014Dokument29 SeitenZhang 2014Saddam FuadNoch keine Bewertungen
- Viapublishedarticle 11 April 2017Dokument7 SeitenViapublishedarticle 11 April 2017Saddam FuadNoch keine Bewertungen
- Intrauterine Fetal TransfusionsDokument10 SeitenIntrauterine Fetal TransfusionsSaddam FuadNoch keine Bewertungen
- Screen-and-Treat Approaches For Cervical Cancer Prevention in Low-Resource SettingsDokument9 SeitenScreen-and-Treat Approaches For Cervical Cancer Prevention in Low-Resource SettingsSaddam FuadNoch keine Bewertungen
- Full-DikonversiDokument9 SeitenFull-DikonversiSaddam FuadNoch keine Bewertungen
- Thrombin 10 PDFDokument5 SeitenThrombin 10 PDFSaddam FuadNoch keine Bewertungen
- PAR in Hemostasis 5Dokument8 SeitenPAR in Hemostasis 5Saddam FuadNoch keine Bewertungen
- 3 13Dokument6 Seiten3 13Muhammad AzkaNoch keine Bewertungen
- Impact of Haemostatic Mechanisms On Pathophysiology of Preeclampsia - 2017 - Thrombosis Research PDFDokument5 SeitenImpact of Haemostatic Mechanisms On Pathophysiology of Preeclampsia - 2017 - Thrombosis Research PDFfujimeisterNoch keine Bewertungen
- Breech 5 PDFDokument2 SeitenBreech 5 PDFSaddam FuadNoch keine Bewertungen
- Roles of Coagulation Proteases and Pars (Protease-Activated Receptors) in Mouse Models of Inflammatory DiseasesDokument12 SeitenRoles of Coagulation Proteases and Pars (Protease-Activated Receptors) in Mouse Models of Inflammatory DiseasesSaddam FuadNoch keine Bewertungen
- Reviews: Pre-Eclampsia Part 2: Prediction, Prevention and ManagementDokument10 SeitenReviews: Pre-Eclampsia Part 2: Prediction, Prevention and Managementdaeng caturNoch keine Bewertungen
- 5 DikonversiDokument14 Seiten5 DikonversiSaddam FuadNoch keine Bewertungen
- Lucena2019 PDFDokument7 SeitenLucena2019 PDFSaddam FuadNoch keine Bewertungen
- Egan2017 DikonversiDokument15 SeitenEgan2017 DikonversiSaddam FuadNoch keine Bewertungen
- Egan 2017Dokument9 SeitenEgan 2017Saddam FuadNoch keine Bewertungen
- 5 DikonversiDokument9 Seiten5 DikonversiSaddam FuadNoch keine Bewertungen
- Lucena2019 PDFDokument7 SeitenLucena2019 PDFSaddam FuadNoch keine Bewertungen
- Aev374 DikonversiDokument14 SeitenAev374 DikonversiSaddam FuadNoch keine Bewertungen
- Paediatrica IndonesianaDokument8 SeitenPaediatrica IndonesianaAnna ClarkNoch keine Bewertungen
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Drug Distribution MethodsDokument40 SeitenDrug Distribution MethodsMuhammad Masoom Akhtar100% (1)
- Lesson I. Background InformationDokument21 SeitenLesson I. Background InformationsuidivoNoch keine Bewertungen
- Soal Midtest + Kunci JawabanDokument28 SeitenSoal Midtest + Kunci JawabanYuyun RasulongNoch keine Bewertungen
- Donna Claire B. Cañeza: Central Bicol State University of AgricultureDokument8 SeitenDonna Claire B. Cañeza: Central Bicol State University of AgricultureDanavie AbergosNoch keine Bewertungen
- DLL Week 5Dokument3 SeitenDLL Week 5Nen CampNoch keine Bewertungen
- Chapter Three: Research MethodologyDokument3 SeitenChapter Three: Research MethodologyEng Abdulkadir MahamedNoch keine Bewertungen
- Thesis Committee MeetingDokument7 SeitenThesis Committee Meetingafknojbcf100% (2)
- A Global StudyDokument57 SeitenA Global StudyRoynal PasaribuNoch keine Bewertungen
- Asset Integrity ManagementDokument5 SeitenAsset Integrity ManagementLuis100% (2)
- Entrepreneur: Job/Career InvolvementDokument5 SeitenEntrepreneur: Job/Career InvolvementYlaissa GeronimoNoch keine Bewertungen
- WWW - Ib.academy: Study GuideDokument122 SeitenWWW - Ib.academy: Study GuideHendrikEspinozaLoyola100% (2)
- 1999, 2003 - Purple Triangles - BrochureDokument32 Seiten1999, 2003 - Purple Triangles - BrochureMaria Patinha100% (2)
- Reflection Paper-The Elephant Man PDFDokument1 SeiteReflection Paper-The Elephant Man PDFCarlosJohn02Noch keine Bewertungen
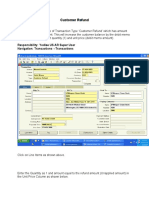
- Customer Refund: Responsibility: Yodlee US AR Super User Navigation: Transactions TransactionsDokument12 SeitenCustomer Refund: Responsibility: Yodlee US AR Super User Navigation: Transactions TransactionsAziz KhanNoch keine Bewertungen
- Class NotesDokument16 SeitenClass NotesAdam AnwarNoch keine Bewertungen
- LakmeDokument34 SeitenLakmeSuraj Pratap Sawhney79% (14)
- Donor S Tax Exam AnswersDokument6 SeitenDonor S Tax Exam AnswersAngela Miles DizonNoch keine Bewertungen
- OrthoDokument22 SeitenOrthosivaleela gNoch keine Bewertungen
- Practice Makes Perfect Basic Spanish Premium Third Edition Dorothy Richmond All ChapterDokument67 SeitenPractice Makes Perfect Basic Spanish Premium Third Edition Dorothy Richmond All Chaptereric.temple792100% (3)
- Congental Abdominal Wall DefectsDokument38 SeitenCongental Abdominal Wall DefectsAhmad Abu KushNoch keine Bewertungen
- Air Augmented Rocket (285pages) Propulsion ConceptsDokument285 SeitenAir Augmented Rocket (285pages) Propulsion ConceptsAlexandre PereiraNoch keine Bewertungen
- Pace, ART 102, Week 6, Etruscan, Roman Arch. & SculpDokument36 SeitenPace, ART 102, Week 6, Etruscan, Roman Arch. & SculpJason ByrdNoch keine Bewertungen
- The ReformationDokument20 SeitenThe ReformationIlyes FerenczNoch keine Bewertungen
- Muller-Lyer IllusionDokument3 SeitenMuller-Lyer Illusionsara VermaNoch keine Bewertungen
- Vrushalirhatwal (14 0)Dokument5 SeitenVrushalirhatwal (14 0)GuruRakshithNoch keine Bewertungen
- Mathematicaleconomics PDFDokument84 SeitenMathematicaleconomics PDFSayyid JifriNoch keine Bewertungen
- Reading İzmir Culture Park Through Women S Experiences Matinee Practices in The 1970s Casino SpacesDokument222 SeitenReading İzmir Culture Park Through Women S Experiences Matinee Practices in The 1970s Casino SpacesAta SagirogluNoch keine Bewertungen
- Entity-Level Controls Fraud QuestionnaireDokument8 SeitenEntity-Level Controls Fraud QuestionnaireKirby C. LoberizaNoch keine Bewertungen
- Names of AllahDokument8 SeitenNames of AllahAfshaan BanuNoch keine Bewertungen
- Tugas, MO - REVIEW JURNAL JIT - Ikomang Aditya Prawira Nugraha (1902612010304)Dokument12 SeitenTugas, MO - REVIEW JURNAL JIT - Ikomang Aditya Prawira Nugraha (1902612010304)MamanxNoch keine Bewertungen