Das könnte Ihnen auch gefallen
- Disorders Potassium 2023-24Dokument18 SeitenDisorders Potassium 2023-24Francisco C. Belloso HornaNoch keine Bewertungen
- 2023 ACC Atrial FibrillationDokument171 Seiten2023 ACC Atrial FibrillationHill kanjinNoch keine Bewertungen
- Diabetic Ketoacidosis 1Dokument10 SeitenDiabetic Ketoacidosis 1Victor Manuel Gonzalez GalindoNoch keine Bewertungen
- Early Versus Late Caffeine Therapy Administration in Preterm Neonates An Updated Systematic Review and Meta-AnalysisDokument10 SeitenEarly Versus Late Caffeine Therapy Administration in Preterm Neonates An Updated Systematic Review and Meta-AnalysisFrancisco C. Belloso HornaNoch keine Bewertungen
- Icu Acquired WeaknessDokument8 SeitenIcu Acquired WeaknessSrinivas PingaliNoch keine Bewertungen
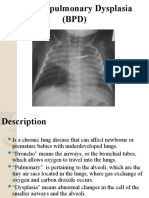
- Bronchopulmonary Dysplasia 2018Dokument20 SeitenBronchopulmonary Dysplasia 2018Francisco C. Belloso HornaNoch keine Bewertungen
- Management of Diabetes Mellitus-Tuberculosis: A Guide To The Essential PracticeDokument99 SeitenManagement of Diabetes Mellitus-Tuberculosis: A Guide To The Essential PracticeFrancisco C. Belloso HornaNoch keine Bewertungen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Wright Et Al 2022 PDFDokument9 SeitenWright Et Al 2022 PDFRadu CiprianNoch keine Bewertungen
- BPD Practice ToolDokument6 SeitenBPD Practice ToolRogelio TorresNoch keine Bewertungen
- Prematurity DR KazevuDokument22 SeitenPrematurity DR KazevuMoses Jr KazevuNoch keine Bewertungen
- Pitfalls, Problems, and Progress in Bronchopulmonary DysplasiaDokument14 SeitenPitfalls, Problems, and Progress in Bronchopulmonary Dysplasiatrixie2k7Noch keine Bewertungen
- 1615 PDFDokument897 Seiten1615 PDFAmmar Mohamad Ziyadi100% (2)
- Pediatrics 2014 Polin 156 63 PDFDokument11 SeitenPediatrics 2014 Polin 156 63 PDFdediraihanNoch keine Bewertungen
- BPDDokument27 SeitenBPDDana GeorgianaNoch keine Bewertungen
- BPD Group 3Dokument13 SeitenBPD Group 3Sheila Rose SalesNoch keine Bewertungen
- Prevention of Bronchopulmonary Dysplasia A Summary of Evidence Based StrategiesDokument15 SeitenPrevention of Bronchopulmonary Dysplasia A Summary of Evidence Based StrategiesMatias LoreiroNoch keine Bewertungen
- BRONCHOPULMONARY DYSPLASIA ModifiedDokument48 SeitenBRONCHOPULMONARY DYSPLASIA ModifiedajanmjNoch keine Bewertungen
- Less Invasive Surfactant Administration: Best Practices and Unanswered QuestionsDokument7 SeitenLess Invasive Surfactant Administration: Best Practices and Unanswered QuestionsRebeca GusmaoNoch keine Bewertungen
- Crack The Core Exam - Volume 1 4th PDFDokument551 SeitenCrack The Core Exam - Volume 1 4th PDFJavier Palma Rojas50% (4)
- Neonatal Respiratory Distress - A Practical Approach To Its Diagnosis and Management 2015Dokument17 SeitenNeonatal Respiratory Distress - A Practical Approach To Its Diagnosis and Management 2015Lissey FloresNoch keine Bewertungen
- Non-Invasive Respiratory Support in Preterm Infants 2022Dokument7 SeitenNon-Invasive Respiratory Support in Preterm Infants 2022DraalexNoch keine Bewertungen
- Guideline For The Prevention of Bronchopulmonary Dysplasia and Assessment of Evolving BPDDokument28 SeitenGuideline For The Prevention of Bronchopulmonary Dysplasia and Assessment of Evolving BPDjanfk2000Noch keine Bewertungen
- (Lung Biology in Health and Disease, V. 137) Richard D Bland - Jacqueline J Coalson-Chronic Lung Disease in Early Infancy-Marcel Dekker (2000)Dokument1.080 Seiten(Lung Biology in Health and Disease, V. 137) Richard D Bland - Jacqueline J Coalson-Chronic Lung Disease in Early Infancy-Marcel Dekker (2000)Uri MoraNoch keine Bewertungen
- Respiratory Support in Preterm Infants at Birth AAP 2014Dokument4 SeitenRespiratory Support in Preterm Infants at Birth AAP 2014Lissey FloresNoch keine Bewertungen
- Follow-Up For NICU Graduates: Howard Needelman Barbara J. JacksonDokument352 SeitenFollow-Up For NICU Graduates: Howard Needelman Barbara J. JacksonStollery SandboxNoch keine Bewertungen
- Non-Invasive High-Frequency Oscillatory Ventilation in Neonates: Review of Physiology, Biology and Clinical DataDokument7 SeitenNon-Invasive High-Frequency Oscillatory Ventilation in Neonates: Review of Physiology, Biology and Clinical DataDaiane BorgesNoch keine Bewertungen
- HAAD Standard For Prophylaxis To Reduce The Risk of RSVDokument4 SeitenHAAD Standard For Prophylaxis To Reduce The Risk of RSVsyedNoch keine Bewertungen
- Maroon and Teal Employee NewsletterDokument4 SeitenMaroon and Teal Employee NewsletterMediception ScienceNoch keine Bewertungen
- Bronchopulmonary DysplasiaDokument17 SeitenBronchopulmonary DysplasiasnphNoch keine Bewertungen
- Bronchopulmonary DysplasiaDokument17 SeitenBronchopulmonary DysplasiaRuDy RaviNoch keine Bewertungen
- Assessment of Postnatal Corticosteroids For The Prevention of BronchopulmonaryDokument20 SeitenAssessment of Postnatal Corticosteroids For The Prevention of Bronchopulmonarylink_wolfloboNoch keine Bewertungen
- Bronchopulmonary Dysplasia (BPD)Dokument13 SeitenBronchopulmonary Dysplasia (BPD)Joan BuiNoch keine Bewertungen
- Treatment PDADokument8 SeitenTreatment PDAijaldo ajahNoch keine Bewertungen
- Pulmonary Manifestations of Pediatric DiseasesDokument356 SeitenPulmonary Manifestations of Pediatric Diseasesmihaela_bondocNoch keine Bewertungen
- 3 Chapter 26 Preterm Postterm NewbornDokument50 Seiten3 Chapter 26 Preterm Postterm NewbornElaiza Anne SanchezNoch keine Bewertungen
- 04 Pediatri Spotleri 2020Dokument104 Seiten04 Pediatri Spotleri 2020Osama E. ShamsNoch keine Bewertungen
- 1193 FullDokument10 Seiten1193 FullzigijuNoch keine Bewertungen