Das könnte Ihnen auch gefallen
- There Is No Doubt ThatDokument7 SeitenThere Is No Doubt ThatAhmad Beni RoufNoch keine Bewertungen
- Reed2014 PDFDokument13 SeitenReed2014 PDFAhmad Beni RoufNoch keine Bewertungen
- Microplate Reader Rayto RT 2100 CDokument1 SeiteMicroplate Reader Rayto RT 2100 CAhmad Beni RoufNoch keine Bewertungen
- I09214649 PDFDokument4 SeitenI09214649 PDFAhmad Beni RoufNoch keine Bewertungen
- Naumowicz2017 Article IntracohortCannibalismAndMethoDokument16 SeitenNaumowicz2017 Article IntracohortCannibalismAndMethoAhmad Beni RoufNoch keine Bewertungen
- Solomon and Udoji PDFDokument14 SeitenSolomon and Udoji PDFAhmad Beni RoufNoch keine Bewertungen
- Soul Lie Re 2017Dokument5 SeitenSoul Lie Re 2017Ahmad Beni RoufNoch keine Bewertungen
- Jurnal Ilmiah Mahasiswa Pendidikan Biologi, Volume 1, Issue 1, Agustus 2016, Hal 95-106Dokument12 SeitenJurnal Ilmiah Mahasiswa Pendidikan Biologi, Volume 1, Issue 1, Agustus 2016, Hal 95-106Ahmad Beni RoufNoch keine Bewertungen
- Brosur ToyoboDokument8 SeitenBrosur ToyobodanilriosNoch keine Bewertungen
- Viral Vaccine For Farmed FishDokument17 SeitenViral Vaccine For Farmed FishAhmad Beni RoufNoch keine Bewertungen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- A Guide To Radio Frequency Hazards With Electric DetonatorsDokument37 SeitenA Guide To Radio Frequency Hazards With Electric Detonatorszix013Noch keine Bewertungen
- MKQ 019Dokument6 SeitenMKQ 019faisaldanyaniNoch keine Bewertungen
- Negative: What Does It Mean To Have A Test Result?Dokument2 SeitenNegative: What Does It Mean To Have A Test Result?Todd EddyNoch keine Bewertungen
- Community Service Synthesis EssayDokument5 SeitenCommunity Service Synthesis Essayapi-343567850Noch keine Bewertungen
- Language Development: - Iphy OkoyeDokument9 SeitenLanguage Development: - Iphy OkoyeIPHY OKOYENoch keine Bewertungen
- Edu101 Debate IsteachingaprofessionDokument3 SeitenEdu101 Debate Isteachingaprofessionapi-236906829Noch keine Bewertungen
- Drug Study TramadolDokument2 SeitenDrug Study TramadolKersey Adricula RicaldeNoch keine Bewertungen
- Microbiome and CancerDokument63 SeitenMicrobiome and CancerK C Goutham ReddyNoch keine Bewertungen
- Gender Differences in AKI To CKD TransitionDokument13 SeitenGender Differences in AKI To CKD TransitionNatasha AlbaShakiraNoch keine Bewertungen
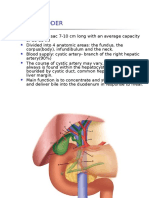
- GALLBLADDERDokument16 SeitenGALLBLADDERKadenceFreya-Charisse G PosadasBulintao100% (2)
- Shell Gadus S3 Wirerope T: Performance, Features & Benefits Main ApplicationsDokument2 SeitenShell Gadus S3 Wirerope T: Performance, Features & Benefits Main ApplicationssfkcrcnNoch keine Bewertungen
- Use of Mosquito Repellent Devices-Problems and Prospects: August 2018Dokument6 SeitenUse of Mosquito Repellent Devices-Problems and Prospects: August 2018Cyrus Ian LanuriasNoch keine Bewertungen
- Introduction To Human BodyDokument57 SeitenIntroduction To Human Bodyjunitria13Noch keine Bewertungen
- Substanceabuseand Trauma: Shannon Simmons,, Liza SuárezDokument12 SeitenSubstanceabuseand Trauma: Shannon Simmons,, Liza SuárezEma UrzicaNoch keine Bewertungen
- Kirloskar Technologies P. LTD.: Appraisal FormDokument4 SeitenKirloskar Technologies P. LTD.: Appraisal Formdihudi.prasantaNoch keine Bewertungen
- Acupuntura y Tui Na en DismenorreaDokument3 SeitenAcupuntura y Tui Na en DismenorreaCristian Dionisio Barros OsorioNoch keine Bewertungen
- Method StatementDokument20 SeitenMethod StatementCharles IkenyeNoch keine Bewertungen
- Nursing 2 4yDokument10 SeitenNursing 2 4yGCON KURNOOLNoch keine Bewertungen
- The Independent 21 February 2016Dokument60 SeitenThe Independent 21 February 2016artedlcNoch keine Bewertungen
- Maintenance Manual: SMV 10-65 B / SMV 37-52 B GANTRYDokument136 SeitenMaintenance Manual: SMV 10-65 B / SMV 37-52 B GANTRYJesùs AlbertoNoch keine Bewertungen
- Past ContinuousDokument2 SeitenPast ContinuousJoaquin GabilondoNoch keine Bewertungen
- Very Important Paper AyurvedaDokument4 SeitenVery Important Paper Ayurvedaarun.sunaniya8701Noch keine Bewertungen
- Tas NeonateDokument13 SeitenTas NeonateAnusha SainiNoch keine Bewertungen
- Vector Borne DiseasesDokument16 SeitenVector Borne Diseasesanon_931122078Noch keine Bewertungen
- Approach To Pateint With Edema Print VersionDokument14 SeitenApproach To Pateint With Edema Print VersionmaybeNoch keine Bewertungen
- Silkpeel DermalInfusion Patient BrochureDokument12 SeitenSilkpeel DermalInfusion Patient BrochureRogier van der HeideNoch keine Bewertungen
- Feu-Nrmf (Drug Study)Dokument7 SeitenFeu-Nrmf (Drug Study)Kaye LaraganNoch keine Bewertungen
- 3rd Quarter Examination in Hope 4Dokument5 Seiten3rd Quarter Examination in Hope 4Hazel Joan Tan100% (3)
- Sample Cbjip Book of Barangay Kauswagan 1Dokument38 SeitenSample Cbjip Book of Barangay Kauswagan 1Johana Pinagayao AngkadNoch keine Bewertungen
- 336 20 006 US Flash Freezing Industrial HempDokument2 Seiten336 20 006 US Flash Freezing Industrial HempJuan Luis Mejias GarciaNoch keine Bewertungen