Das könnte Ihnen auch gefallen
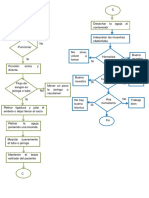
- Diagrama de Flujo Muestra SangreDokument2 SeitenDiagrama de Flujo Muestra SangreSecc. 2 Marco Antonio Aviles Romero100% (4)
- Diagrama de Flujo Muestra SangreDokument2 SeitenDiagrama de Flujo Muestra SangreSecc. 2 Marco Antonio Aviles Romero100% (4)
- Cultivos HidroponicosDokument56 SeitenCultivos HidroponicosAndres F Patiño Sanchez100% (3)
- Ciencias I Biologia Planeacion Bimestral I 2011 2012Dokument12 SeitenCiencias I Biologia Planeacion Bimestral I 2011 2012Profesor de Biología94% (17)
- Primer Año Bachillerato Biologia 2018Dokument19 SeitenPrimer Año Bachillerato Biologia 2018Sandra Quimi-Cruz100% (1)
- Ramos Perez Silverstone Sopkin 2018 Catalogo de La Flora Relictual OcrDokument238 SeitenRamos Perez Silverstone Sopkin 2018 Catalogo de La Flora Relictual Ocree m (Elsoad)Noch keine Bewertungen
- Proyecto Fisica para Arquitectura: Techos VerdesDokument26 SeitenProyecto Fisica para Arquitectura: Techos VerdesLuciana AguilarNoch keine Bewertungen
- NOTA: 92 Les Dije Que No Hicieran PortadaDokument5 SeitenNOTA: 92 Les Dije Que No Hicieran PortadaRicardo Barboza MontealegreNoch keine Bewertungen
- Ciclo Del FósforoDokument12 SeitenCiclo Del FósforoSecc. 2 Marco Antonio Aviles RomeroNoch keine Bewertungen
- Aterosclerosis Coronaria y Daño A MiocardioDokument9 SeitenAterosclerosis Coronaria y Daño A MiocardioSecc. 2 Marco Antonio Aviles RomeroNoch keine Bewertungen
- Ciclo Del FósforoDokument12 SeitenCiclo Del FósforoSecc. 2 Marco Antonio Aviles RomeroNoch keine Bewertungen
- Ciclo Del NitrógenoDokument29 SeitenCiclo Del NitrógenoSecc. 2 Marco Antonio Aviles RomeroNoch keine Bewertungen
- Info Agro TEXTURA ....Dokument5 SeitenInfo Agro TEXTURA ....DavidJ.MarchanGuerreroNoch keine Bewertungen
- Ciclo Del AzufreDokument16 SeitenCiclo Del AzufreSecc. 2 Marco Antonio Aviles RomeroNoch keine Bewertungen
- EspectrofotometriaDokument2 SeitenEspectrofotometriaSecc. 2 Marco Antonio Aviles RomeroNoch keine Bewertungen
- Examensito Suelitos PHDokument1 SeiteExamensito Suelitos PHSecc. 2 Marco Antonio Aviles RomeroNoch keine Bewertungen
- Informacion IgEDokument4 SeitenInformacion IgESecc. 2 Marco Antonio Aviles RomeroNoch keine Bewertungen
- Phmetro Mesa y PortatilDokument2 SeitenPhmetro Mesa y PortatilSecc. 2 Marco Antonio Aviles RomeroNoch keine Bewertungen
- Informacion IgEDokument4 SeitenInformacion IgESecc. 2 Marco Antonio Aviles RomeroNoch keine Bewertungen
- EspectrofotometriaDokument2 SeitenEspectrofotometriaSecc. 2 Marco Antonio Aviles RomeroNoch keine Bewertungen
- Aminoacidos EsencialesDokument6 SeitenAminoacidos EsencialesSecc. 2 Marco Antonio Aviles RomeroNoch keine Bewertungen
- P2 5QM1 AgoDic2019Dokument1 SeiteP2 5QM1 AgoDic2019Secc. 2 Marco Antonio Aviles RomeroNoch keine Bewertungen
- Efecto Citopatico VirusDokument1 SeiteEfecto Citopatico VirusSecc. 2 Marco Antonio Aviles RomeroNoch keine Bewertungen
- Extraccion de MuestraDokument5 SeitenExtraccion de MuestraBrenda SantosNoch keine Bewertungen
- Bacterial and Bacteriophage GeneticsDokument8 SeitenBacterial and Bacteriophage GeneticsBetty LopezNoch keine Bewertungen
- Preguntas FisiologiaDokument2 SeitenPreguntas FisiologiaSecc. 2 Marco Antonio Aviles RomeroNoch keine Bewertungen
- Catorce Puntos de Deming PDFDokument26 SeitenCatorce Puntos de Deming PDFSecc. 2 Marco Antonio Aviles RomeroNoch keine Bewertungen
- InformeabejasinviernoDokument17 SeitenInformeabejasinviernoValentinaAlarcónNoch keine Bewertungen
- Tipos de COX HumanasDokument2 SeitenTipos de COX HumanasSecc. 2 Marco Antonio Aviles RomeroNoch keine Bewertungen
- Efecto Citopatico VirusDokument1 SeiteEfecto Citopatico VirusSecc. 2 Marco Antonio Aviles RomeroNoch keine Bewertungen
- Hidroponia y HumusDokument12 SeitenHidroponia y HumusEmilio Cordova ChevalierNoch keine Bewertungen
- Taller 1 - Taxonomía y ReinosDokument3 SeitenTaller 1 - Taxonomía y ReinosKatherineParraNoch keine Bewertungen
- Texto Biologia 2020Dokument91 SeitenTexto Biologia 2020Henrry CondoriNoch keine Bewertungen
- Guia Nat 2°Dokument2 SeitenGuia Nat 2°FERNANDANoch keine Bewertungen
- Some QuestionsDokument5 SeitenSome QuestionsJuan Camiloo Quinteroo VargasNoch keine Bewertungen
- 1er Grado Abril - Instrumentos de Evaluación (2023-2024)Dokument13 Seiten1er Grado Abril - Instrumentos de Evaluación (2023-2024)mateoeguia3119Noch keine Bewertungen
- Biología, Geología y Ciencias AmbientalesDokument15 SeitenBiología, Geología y Ciencias AmbientalesJAVIER GARCIA CALLEJANoch keine Bewertungen
- Saliva y ElodeaDokument2 SeitenSaliva y ElodeaGerson LopezNoch keine Bewertungen
- Resumen Biologia para Bachillerato PDFDokument116 SeitenResumen Biologia para Bachillerato PDFAlex OlmosNoch keine Bewertungen
- Influencia Del Tipo de Luz en Plántulas de Frijol (Phaseolus Vulgaris)Dokument14 SeitenInfluencia Del Tipo de Luz en Plántulas de Frijol (Phaseolus Vulgaris)Jesus UscangaNoch keine Bewertungen
- Guía Rápida CBS UnlockedDokument7 SeitenGuía Rápida CBS UnlockedGuia MateriasNoch keine Bewertungen
- Como Nace La MicrobiologiaDokument12 SeitenComo Nace La Microbiologiavictor mendoza0% (1)
- Modulo de Ciencias Del Trigo Al PanDokument65 SeitenModulo de Ciencias Del Trigo Al PanSulvy Cáceres VezgaNoch keine Bewertungen
- Señales Luminosas y Desarrollo de Las PlantasDokument4 SeitenSeñales Luminosas y Desarrollo de Las PlantasGuille AlvarengaNoch keine Bewertungen
- Ensayo BotanicaDokument2 SeitenEnsayo BotanicaRoque Guevara ArrietaNoch keine Bewertungen
- Natu 4 Doc AnimateDokument48 SeitenNatu 4 Doc AnimateAlejandra FloresNoch keine Bewertungen
- TRABAJO de INVESTIGACIÓN (Actividad Antifúngica de Compuestos Fenólicos de La Tara Frente Al Fusarium Graminearum)Dokument23 SeitenTRABAJO de INVESTIGACIÓN (Actividad Antifúngica de Compuestos Fenólicos de La Tara Frente Al Fusarium Graminearum)Melissa HernandezNoch keine Bewertungen
- TEXTO-PARALELO-fundamento de Ciencias Naturalez y Del Medio AmbienteDokument51 SeitenTEXTO-PARALELO-fundamento de Ciencias Naturalez y Del Medio AmbienteElenaNoch keine Bewertungen
- Solucion Del Taller de AgropecuariaDokument19 SeitenSolucion Del Taller de Agropecuariajazmin camachoNoch keine Bewertungen
- Guia 7,8,9 Ciencias Naturales 5° Básico NEEP ListoDokument19 SeitenGuia 7,8,9 Ciencias Naturales 5° Básico NEEP ListoNicole FernandezNoch keine Bewertungen
- Estres Hidrico en Limon TahitiDokument5 SeitenEstres Hidrico en Limon TahitiPaul Poma GuevaraNoch keine Bewertungen
- Biologa Cuarto AoDokument130 SeitenBiologa Cuarto AoseemmartNoch keine Bewertungen
- Papa InformeDokument3 SeitenPapa InformeLukaz ReyesNoch keine Bewertungen
- Guia de Uso de Charolas Germinadoras PDFDokument6 SeitenGuia de Uso de Charolas Germinadoras PDFWilder Castillo VelásquezNoch keine Bewertungen