Das könnte Ihnen auch gefallen
- El cannabis en patologías del sistema nervioso centralVon EverandEl cannabis en patologías del sistema nervioso centralNoch keine Bewertungen
- Ensayo de Psicologia - 245Dokument9 SeitenEnsayo de Psicologia - 245Alanoca MamaniNoch keine Bewertungen
- Aprenda Farmacología En Una SemanaVon EverandAprenda Farmacología En Una SemanaBewertung: 2 von 5 Sternen2/5 (1)
- Seminario de NeurotransmisoresDokument3 SeitenSeminario de NeurotransmisoresMariafé TaboadaNoch keine Bewertungen
- NeurotransmisoresDokument4 SeitenNeurotransmisoresPaula TarapuesNoch keine Bewertungen
- Neurotransmisores 40Dokument11 SeitenNeurotransmisores 40bella esmeralda carbajal mayhuaNoch keine Bewertungen
- NeurotransmisoresDokument2 SeitenNeurotransmisoresJoseth Carolina SantanaNoch keine Bewertungen
- Cate Cola MinasDokument24 SeitenCate Cola MinasRoman Ortega67% (3)
- Neurotransmisores claveDokument28 SeitenNeurotransmisores claveEloíss G. Palacios GallegoNoch keine Bewertungen
- Neurotransmisores pequeños y rápidosDokument28 SeitenNeurotransmisores pequeños y rápidosAndreaNoch keine Bewertungen
- Neuro Receptor EsDokument3 SeitenNeuro Receptor EsThomas StetickNoch keine Bewertungen
- Aminoácidos Como Neurotransmisores-BioquimicaDokument4 SeitenAminoácidos Como Neurotransmisores-BioquimicaCedeño González Lucero EsmeraldaNoch keine Bewertungen
- Los Neurotransmisores Más ImportantesDokument2 SeitenLos Neurotransmisores Más ImportantesTania CallizanaNoch keine Bewertungen
- Morfologia SinapticaDokument4 SeitenMorfologia SinapticaIvan Figueroa MartinezNoch keine Bewertungen
- Neurotransmisores Doc BaseDokument13 SeitenNeurotransmisores Doc BasePetra PérezNoch keine Bewertungen
- Bases Biologicas Renzo Pulache PantaDokument5 SeitenBases Biologicas Renzo Pulache Pantajaime eduardoNoch keine Bewertungen
- 2.5.1 VÍAS SEROTONINÉRGICAS, DOPAMINÉRGICAS Y DOPAMINÉRGICASy NoradrenérgicaDokument4 Seiten2.5.1 VÍAS SEROTONINÉRGICAS, DOPAMINÉRGICAS Y DOPAMINÉRGICASy NoradrenérgicaSofía CelisNoch keine Bewertungen
- Resumen NeurotransmisoresDokument4 SeitenResumen NeurotransmisoresHéctor CervantesNoch keine Bewertungen
- FarmacologiaDokument21 SeitenFarmacologiajorgeosmarr33% (3)
- Catecolaminas 120815191953 Phpapp01Dokument24 SeitenCatecolaminas 120815191953 Phpapp01elieliabregoNoch keine Bewertungen
- Neurotransmisores y ClasificacionDokument59 SeitenNeurotransmisores y ClasificacionCeci Flores100% (1)
- Resumen de NeurotransmisoresDokument4 SeitenResumen de NeurotransmisoresBelial Bolaños100% (1)
- Fármacos Que Actúan en El SNADokument4 SeitenFármacos Que Actúan en El SNAceciiliiaa_rNoch keine Bewertungen
- Sem1 Trabajo de NeurotransmisoresDokument19 SeitenSem1 Trabajo de NeurotransmisoresFranco RomeroNoch keine Bewertungen
- Wa0016.Dokument17 SeitenWa0016.gomez2006lauNoch keine Bewertungen
- Conferencia 8Dokument17 SeitenConferencia 8David CampoverdeNoch keine Bewertungen
- 1.2.1.1. Adrenalina y NoradrenalinaDokument11 Seiten1.2.1.1. Adrenalina y NoradrenalinaEd ChávezNoch keine Bewertungen
- Los principales neurotransmisores enDokument18 SeitenLos principales neurotransmisores enSilvia RoncalNoch keine Bewertungen
- Neurotransmisores Fisiologia Animal.Dokument16 SeitenNeurotransmisores Fisiologia Animal.Carlos Daniel Quispe ChuraNoch keine Bewertungen
- TEMA 3.4 BiologiaDokument9 SeitenTEMA 3.4 BiologiaMarta Moron poloNoch keine Bewertungen
- Orientadora 3Dokument12 SeitenOrientadora 3Adri LópezNoch keine Bewertungen
- Ricardo Espinosa 8-1008-1897Dokument6 SeitenRicardo Espinosa 8-1008-1897Nathanael CooteNoch keine Bewertungen
- Principales NeurotransmisoresDokument8 SeitenPrincipales NeurotransmisoresMinttua DaNoch keine Bewertungen
- Cuestiuonerio 2Dokument31 SeitenCuestiuonerio 2Melanie Castro100% (1)
- Seminario 03 CompletoDokument19 SeitenSeminario 03 CompletoReynaldo FaggianiNoch keine Bewertungen
- Taller Histofisiolofia Del Sistema Nervioso y NeurotransmisoresDokument4 SeitenTaller Histofisiolofia Del Sistema Nervioso y NeurotransmisoresAngie Nathalia Cutiva MedinaNoch keine Bewertungen
- 6 - Teoría - Vereau - Neurotransmisores PDFDokument37 Seiten6 - Teoría - Vereau - Neurotransmisores PDFSol CamusNoch keine Bewertungen
- Importancia Clinica de La Farmacologia de Los Receptores y Sistema Nervioso AutonomoDokument17 SeitenImportancia Clinica de La Farmacologia de Los Receptores y Sistema Nervioso AutonomoArturo Praeli100% (1)
- Neurotransmisores y sinapsisDokument29 SeitenNeurotransmisores y sinapsisCHRISTIAN MORALESNoch keine Bewertungen
- Farm. General - Ao N⺠3Dokument13 SeitenFarm. General - Ao N⺠3Wilder CanelonNoch keine Bewertungen
- NeurotransmisoresDokument30 SeitenNeurotransmisoresAmni JF100% (1)
- Neuro UNIDAD 3Dokument8 SeitenNeuro UNIDAD 3Gabi CasasNoch keine Bewertungen
- 3 Resumen de Farmaco LDokument7 Seiten3 Resumen de Farmaco LsussamvarelaNoch keine Bewertungen
- 212K23074 EN6TV1 Act2Dokument8 Seiten212K23074 EN6TV1 Act2Litzy Raquel Cruz OrtizNoch keine Bewertungen
- Tema 13 (Sinapsis 2)Dokument6 SeitenTema 13 (Sinapsis 2)jesus sanchez coelhoNoch keine Bewertungen
- Farmacología SNCDokument28 SeitenFarmacología SNCevelynNoch keine Bewertungen
- Regulacion de Principios NeutrasmisoresDokument6 SeitenRegulacion de Principios NeutrasmisoresJahaira Lezama GarciaNoch keine Bewertungen
- Eulalia Fernández Vallìn Cárdenas, MDDokument89 SeitenEulalia Fernández Vallìn Cárdenas, MDFernandaNoch keine Bewertungen
- Farmacologia Adrenergicos Parte 1Dokument13 SeitenFarmacologia Adrenergicos Parte 1Wilianny RomeroNoch keine Bewertungen
- RESUMEN Seminario 4Dokument10 SeitenRESUMEN Seminario 4LAUFERNoch keine Bewertungen
- Funcionamiento del cerebro: principales neurotransmisoresDokument17 SeitenFuncionamiento del cerebro: principales neurotransmisoresReyna VaherNoch keine Bewertungen
- Tarea 4Dokument2 SeitenTarea 4Rudy E. FerrerasNoch keine Bewertungen
- Fisiología Seminario 6 Neurotransmisores y NeuropeptidosDokument8 SeitenFisiología Seminario 6 Neurotransmisores y Neuropeptidosuser1818Noch keine Bewertungen
- Anatomía y Fisiología IIDokument28 SeitenAnatomía y Fisiología IIDaniela Solorzano GonzalezNoch keine Bewertungen
- CAPITULO 3 - Neurotransmisores y ReceptoresDokument20 SeitenCAPITULO 3 - Neurotransmisores y ReceptoresParaísos PerdidosNoch keine Bewertungen
- Anatomia y Filosofia Del Sistema-Nervioso Tarea IIDokument15 SeitenAnatomia y Filosofia Del Sistema-Nervioso Tarea IIurielNoch keine Bewertungen
- UNIVERSIDAD NACIONAL AUTÓNOMA DE HONDURAS EN EL VALLE DE SULADokument82 SeitenUNIVERSIDAD NACIONAL AUTÓNOMA DE HONDURAS EN EL VALLE DE SULALarolyn RodriguezNoch keine Bewertungen
- Fármacos SNA: neurotransmisores SNADokument4 SeitenFármacos SNA: neurotransmisores SNArodrigo4590Noch keine Bewertungen
- Seminario II - Cuestionario 2Dokument3 SeitenSeminario II - Cuestionario 2usmpusmpusmpNoch keine Bewertungen
- Las Moléculas de Los NeurotransmisoresDokument4 SeitenLas Moléculas de Los NeurotransmisoresAntonio LeónNoch keine Bewertungen
- Actividad 5 y 6Dokument7 SeitenActividad 5 y 6Cecilia Enciso MontejoNoch keine Bewertungen
- NeoplasiasDokument22 SeitenNeoplasiasCecilia Enciso MontejoNoch keine Bewertungen
- Trabajo Final - Evolucion GeologicaDokument5 SeitenTrabajo Final - Evolucion GeologicaCecilia Enciso MontejoNoch keine Bewertungen
- Tarea de Análisis ClínicoDokument1 SeiteTarea de Análisis ClínicoCecilia Enciso MontejoNoch keine Bewertungen
- Proyecto EstadisticaDokument4 SeitenProyecto EstadisticaCecilia Enciso MontejoNoch keine Bewertungen
- SindromeDokument10 SeitenSindromeCecilia Enciso MontejoNoch keine Bewertungen
- PX Femenina de 12 Años de EdadDokument2 SeitenPX Femenina de 12 Años de EdadCecilia Enciso MontejoNoch keine Bewertungen
- Preguntas Yas y AngelDokument4 SeitenPreguntas Yas y AngelCecilia Enciso MontejoNoch keine Bewertungen
- Jerarquia Por PosicionDokument7 SeitenJerarquia Por PosicionCecilia Enciso MontejoNoch keine Bewertungen
- Caso ClínicoDokument1 SeiteCaso ClínicoCecilia Enciso MontejoNoch keine Bewertungen
- Exploración de Los Pares CranealesDokument23 SeitenExploración de Los Pares CranealesCecilia Enciso MontejoNoch keine Bewertungen
- Sindrome deDokument8 SeitenSindrome deCecilia Enciso MontejoNoch keine Bewertungen
- Sustancias en El CigarroDokument4 SeitenSustancias en El CigarroCecilia Enciso MontejoNoch keine Bewertungen
- FoliculitisDokument14 SeitenFoliculitisCecilia Enciso MontejoNoch keine Bewertungen
- Ensayo CeciDokument2 SeitenEnsayo CeciCecilia Enciso MontejoNoch keine Bewertungen
- Nota Inicial (Ejemplo)Dokument3 SeitenNota Inicial (Ejemplo)Cecilia Enciso MontejoNoch keine Bewertungen
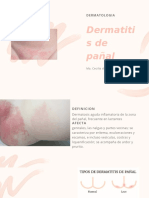
- Dermatitis de pañal: causas, síntomas y tratamientoDokument8 SeitenDermatitis de pañal: causas, síntomas y tratamientoCecilia Enciso MontejoNoch keine Bewertungen
- Síndrome extrapiramidal y ParkinsonDokument59 SeitenSíndrome extrapiramidal y ParkinsonCecilia Enciso MontejoNoch keine Bewertungen
- Síndrome extrapiramidal y ParkinsonDokument59 SeitenSíndrome extrapiramidal y ParkinsonCecilia Enciso MontejoNoch keine Bewertungen
- SR Menopausia ClimaterioDokument48 SeitenSR Menopausia ClimaterioCecilia Enciso MontejoNoch keine Bewertungen
- COLELITIASIS Y SUS COMPLICACIONES Pregrado 2007Dokument67 SeitenCOLELITIASIS Y SUS COMPLICACIONES Pregrado 2007Jorge SumbaNoch keine Bewertungen
- 01.b. Taller de Búsqueda CientíficaDokument37 Seiten01.b. Taller de Búsqueda CientíficaSebastian MonteroNoch keine Bewertungen
- Salud TrabajadoresDokument4 SeitenSalud TrabajadoresAngie GómezNoch keine Bewertungen
- Como Usar Reiki Y Cristales de CuarzoDokument5 SeitenComo Usar Reiki Y Cristales de CuarzoJasmin Torres GodoyNoch keine Bewertungen
- Logoterapia y FenomenologíaDokument6 SeitenLogoterapia y Fenomenologíajuan dNoch keine Bewertungen
- Infografia TeaDokument1 SeiteInfografia TeaViani Sulemm Bello EspinozaNoch keine Bewertungen
- Seminario de Habilidades Directivas Taller de Trabajo N° 7Dokument52 SeitenSeminario de Habilidades Directivas Taller de Trabajo N° 7Ricardo MacrobioNoch keine Bewertungen
- Marco Teorico-. CytDokument6 SeitenMarco Teorico-. CytAndrea TorresNoch keine Bewertungen
- POE 1 Recepción G02Dokument11 SeitenPOE 1 Recepción G02Santos mariela Ramos espinozaNoch keine Bewertungen
- Proyecto Hábitos Alimentacios en Niños y Adolescentes Del GCCDokument40 SeitenProyecto Hábitos Alimentacios en Niños y Adolescentes Del GCCag5031785Noch keine Bewertungen
- BOK-BU: Masaje Abdominal Coreano: Los Beneficios Más Destacados de Esta Técnica Son Los SiguientesDokument3 SeitenBOK-BU: Masaje Abdominal Coreano: Los Beneficios Más Destacados de Esta Técnica Son Los SiguientesGuacimara MedinaNoch keine Bewertungen
- Desarrollo de Sesiones de TutoríaDokument5 SeitenDesarrollo de Sesiones de TutoríaCaridad Plasencia HuamanNoch keine Bewertungen
- Carnet Estacion de NaranjalDokument21 SeitenCarnet Estacion de NaranjalMari GonzalesNoch keine Bewertungen
- Dimensión AfectivaDokument1 SeiteDimensión AfectivaJuan ArizaNoch keine Bewertungen
- Adquisición de La InformaciónDokument5 SeitenAdquisición de La InformaciónJose A ManzaneroNoch keine Bewertungen
- Proyecto de Cria y Comercialización de Carne de Venado Cola Blanca PDFDokument115 SeitenProyecto de Cria y Comercialización de Carne de Venado Cola Blanca PDFVictor Pinelo50% (2)
- Ayuno Intermitente - Como Ayunar de La Mejor Manera CerascreenDokument1 SeiteAyuno Intermitente - Como Ayunar de La Mejor Manera CerascreenromeroandradeximenaNoch keine Bewertungen
- NeostigminaDokument4 SeitenNeostigminaAbel TintaNoch keine Bewertungen
- Taller Reconociendo Mis Propios Riesgos en SMDokument1 SeiteTaller Reconociendo Mis Propios Riesgos en SMFeer Zagarra100% (1)
- AMNANESISDokument5 SeitenAMNANESISTahit TarazonaNoch keine Bewertungen
- Hernia UmbilicalDokument4 SeitenHernia UmbilicalMishu NaranjoNoch keine Bewertungen
- 4Dokument5 Seiten4DavidNoch keine Bewertungen
- Beneficios de La Almendra y Aceite de AlmendraDokument9 SeitenBeneficios de La Almendra y Aceite de AlmendraDaniela Jaqueline Aramayo HinojosaNoch keine Bewertungen
- CS. DE LA SALUD 3° A Análisis e Interpretación EstresDokument2 SeitenCS. DE LA SALUD 3° A Análisis e Interpretación EstresVale Castillo GrandónNoch keine Bewertungen
- Introducción A Los Modelos de Terapia Familiar 2019Dokument107 SeitenIntroducción A Los Modelos de Terapia Familiar 2019Macarena Rodriguez100% (1)
- Reparación colector primario alcantarillado Olmos LambayequeDokument6 SeitenReparación colector primario alcantarillado Olmos LambayequeRonal Ivan Vasquez ZorrillaNoch keine Bewertungen
- g6.pp Guia Tecnica Del Componente de Alimentacion Nutricion Icbf v5 PDFDokument184 Seiteng6.pp Guia Tecnica Del Componente de Alimentacion Nutricion Icbf v5 PDFTaTianaMoraLesNoch keine Bewertungen
- Plantas bioindicadoras ambientalesDokument10 SeitenPlantas bioindicadoras ambientalesJOSÉ DAVID OSPINA TORONoch keine Bewertungen
- Teoria MaslowDokument8 SeitenTeoria MaslowConstructores De OrienteNoch keine Bewertungen
- TEFIS BallenBallenNickH 2014Dokument78 SeitenTEFIS BallenBallenNickH 2014Cesar DiazNoch keine Bewertungen
- Cuadro Comparativo Sorgo y AvenaDokument8 SeitenCuadro Comparativo Sorgo y AvenaLupita GilNoch keine Bewertungen