Das könnte Ihnen auch gefallen
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Vacuum Systems HandbookDokument121 SeitenVacuum Systems Handbookapi-3812573100% (6)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- HAZOP Training290620Dokument93 SeitenHAZOP Training290620NasrulNoch keine Bewertungen
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- Introduction To Storage TanksDokument49 SeitenIntroduction To Storage TanksMachineryeng100% (2)
- IIW - International Institute of WeldingDokument3 SeitenIIW - International Institute of WeldingNilesh MistryNoch keine Bewertungen
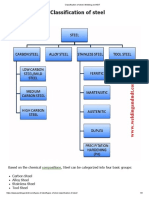
- Classification of Steel - Welding and NDTDokument3 SeitenClassification of Steel - Welding and NDTAshif Iqubal100% (1)
- 9 Ion ExchangeDokument68 Seiten9 Ion ExchangeThiago CangussuNoch keine Bewertungen
- Lesson 1 - Reactions Energy WorksheetDokument4 SeitenLesson 1 - Reactions Energy WorksheetAvakoalaNoch keine Bewertungen
- Coffee Roast ProcessDokument44 SeitenCoffee Roast ProcesstheblueserenadeNoch keine Bewertungen
- Metal CastingDokument40 SeitenMetal CastingFahmi Sanji AlexanderNoch keine Bewertungen
- Coffee RoastingDokument187 SeitenCoffee RoastingAntonio Parlange100% (11)
- Theory To Practice Oxygen Transfer and The New ASCE StandardDokument15 SeitenTheory To Practice Oxygen Transfer and The New ASCE StandardArtesira YunaNoch keine Bewertungen
- PL IS enDokument882 SeitenPL IS enOssian89Noch keine Bewertungen
- Planning Fundamentals IndustryDokument846 SeitenPlanning Fundamentals IndustrymaniyarasanNoch keine Bewertungen
- PL BT enDokument304 SeitenPL BT enOssian89Noch keine Bewertungen
- Pregun Csaba - Water Resources Management and Water Quality ProtectionDokument117 SeitenPregun Csaba - Water Resources Management and Water Quality ProtectionOssian89Noch keine Bewertungen
- Witte - Liquid-Gas Ejektor Design PDFDokument114 SeitenWitte - Liquid-Gas Ejektor Design PDFOssian89Noch keine Bewertungen
- Continuous Removal of AmmoniumIon by IonexchangeDokument8 SeitenContinuous Removal of AmmoniumIon by IonexchangeOssian89Noch keine Bewertungen
- R. Sander: Compilation of Henry's Law ConstantsDokument583 SeitenR. Sander: Compilation of Henry's Law Constantshrvoje_lisacNoch keine Bewertungen
- Bacterial Biofilms in Diabetic Foot UlcersDokument20 SeitenBacterial Biofilms in Diabetic Foot UlcersOssian89Noch keine Bewertungen
- Witte - Liquid-Gas Ejektor Design PDFDokument114 SeitenWitte - Liquid-Gas Ejektor Design PDFOssian89Noch keine Bewertungen
- XAD4 Resin DatasheetDokument4 SeitenXAD4 Resin DatasheetOssian89Noch keine Bewertungen
- Reverse Osmosis Filtration For Space Mission Wastewater PDFDokument14 SeitenReverse Osmosis Filtration For Space Mission Wastewater PDFOssian89Noch keine Bewertungen
- Side Effects of ChlorinationDokument286 SeitenSide Effects of ChlorinationOssian89Noch keine Bewertungen
- Heating On Composition and Thermo-Oxidative Stability of ArabicaDokument13 SeitenHeating On Composition and Thermo-Oxidative Stability of ArabicaOssian89Noch keine Bewertungen
- Quality Enhancement of Coffee With Acid and Enzyme TreatmentDokument131 SeitenQuality Enhancement of Coffee With Acid and Enzyme TreatmentOssian89100% (1)
- Best Practices For Small Drinking Water Systems-ManualDokument51 SeitenBest Practices For Small Drinking Water Systems-ManualSlobodan JankovicNoch keine Bewertungen
- 308 794 1 PBDokument15 Seiten308 794 1 PBDimas RizkyNoch keine Bewertungen
- Efficiency of Domestic Reverse Osmosis in Removal of THMDokument6 SeitenEfficiency of Domestic Reverse Osmosis in Removal of THMOssian89Noch keine Bewertungen
- Design of A Small Scale Iron and Manganese Removal System For Copperbelt Universitys Borehole Water PDFDokument7 SeitenDesign of A Small Scale Iron and Manganese Removal System For Copperbelt Universitys Borehole Water PDFOssian89Noch keine Bewertungen
- Commissioningofwastewatertreatmentplants PDFDokument79 SeitenCommissioningofwastewatertreatmentplants PDFOssian89Noch keine Bewertungen
- Side Effects of ChlorinationDokument286 SeitenSide Effects of ChlorinationOssian89Noch keine Bewertungen
- Alternative Processes For Removal of Organohalogens PDFDokument201 SeitenAlternative Processes For Removal of Organohalogens PDFOssian89100% (1)
- Water Filters Can Effectively Remove Disinfection By-ProductsDokument10 SeitenWater Filters Can Effectively Remove Disinfection By-ProductsOssian89Noch keine Bewertungen
- Design of Municipal Diatomite Filters For Iron RemovalDokument185 SeitenDesign of Municipal Diatomite Filters For Iron RemovalOssian89Noch keine Bewertungen
- EPA - Design Handbook For Automation of As WWTPDokument427 SeitenEPA - Design Handbook For Automation of As WWTPOssian89Noch keine Bewertungen
- Efficient Synthesis of 3-Hydroxy-1,4-Benzodiazepines Oxazepam and Lorazepam by New Acetoxylation Reaction of 3-Position of 1,4-Benzodiazepine Ring - Organic Process Research & DevelopmentDokument12 SeitenEfficient Synthesis of 3-Hydroxy-1,4-Benzodiazepines Oxazepam and Lorazepam by New Acetoxylation Reaction of 3-Position of 1,4-Benzodiazepine Ring - Organic Process Research & DevelopmentSimon GeschwindNoch keine Bewertungen
- Rdna Prac 4Dokument2 SeitenRdna Prac 4Mthetheleli NxeleNoch keine Bewertungen
- International Safety Standards Reference Guide: For The Juvenile Products IndustryDokument32 SeitenInternational Safety Standards Reference Guide: For The Juvenile Products IndustryMaria SilberNoch keine Bewertungen
- Physical and Chemical Properties QuizDokument20 SeitenPhysical and Chemical Properties Quizgnanasekar0% (1)
- A Seminar Report OnDokument1 SeiteA Seminar Report Onu10ch019Noch keine Bewertungen
- ATCO Gas STOPAQ-Vinylester Procedure Rev0Dokument45 SeitenATCO Gas STOPAQ-Vinylester Procedure Rev0mkash028Noch keine Bewertungen
- T50 300 Xl092grexbDokument3 SeitenT50 300 Xl092grexbSykat ZamanNoch keine Bewertungen
- Cuellar Bermudez S P Aguilar Hernandez I PDFDokument20 SeitenCuellar Bermudez S P Aguilar Hernandez I PDFJader MendozaNoch keine Bewertungen
- Interpretation Guide: Aerobic Count PlateDokument6 SeitenInterpretation Guide: Aerobic Count PlateJinalNoch keine Bewertungen
- Wastewater treatment and sludge digestion process explainedDokument17 SeitenWastewater treatment and sludge digestion process explainedParas KathvadiyaNoch keine Bewertungen
- Ch11EN Fluid Unit Flow PathDokument1 SeiteCh11EN Fluid Unit Flow PathHermeson SantiagoNoch keine Bewertungen
- 10.1016/j.foodres.2014.01.057: Food Research InternationalDokument90 Seiten10.1016/j.foodres.2014.01.057: Food Research Internationaledywiyono2013Noch keine Bewertungen
- CH 5 Heat Exchanger Design MethodsDokument30 SeitenCH 5 Heat Exchanger Design Methodsapi-3765936100% (1)
- Life's Origin TheoriesDokument8 SeitenLife's Origin TheoriesSazzad NiloyNoch keine Bewertungen
- 4013 Stability TestingDokument5 Seiten4013 Stability TestingtghonsNoch keine Bewertungen
- Hegatec VacuumBeltDryerDokument2 SeitenHegatec VacuumBeltDryerpintaratNoch keine Bewertungen
- Springwire Xinhua 2006Dokument10 SeitenSpringwire Xinhua 2006teban09Noch keine Bewertungen
- Optek TOP5 Brewery Applications EnglishDokument16 SeitenOptek TOP5 Brewery Applications EnglishoptekNoch keine Bewertungen
- Hot Fill Processing of BeveragesDokument3 SeitenHot Fill Processing of BeveragesJavier Ignacio Leyton SotoNoch keine Bewertungen
- Cambridge IGCSE (9-1) : PHYSICS 0972/41Dokument16 SeitenCambridge IGCSE (9-1) : PHYSICS 0972/41dowanahamidNoch keine Bewertungen
- M MLV Reverse Transcriptase ProtocolDokument2 SeitenM MLV Reverse Transcriptase ProtocolSergio HernandezNoch keine Bewertungen
- E.O.Paton Electric Welding Institute Activity in The FieldDokument10 SeitenE.O.Paton Electric Welding Institute Activity in The FieldMaksimovNoch keine Bewertungen
- AVENDAÑO, Jay Russell A. Written Report PURUGGANAN, Stephanie Claire SISON, KellyDokument4 SeitenAVENDAÑO, Jay Russell A. Written Report PURUGGANAN, Stephanie Claire SISON, KellyKelly SisonNoch keine Bewertungen
- Test Report E-Liquid: 1 Sample InformationDokument10 SeitenTest Report E-Liquid: 1 Sample InformationVangel PetrovNoch keine Bewertungen