Das könnte Ihnen auch gefallen
- Imunossenescência: envelhecimento do sistema imuneVon EverandImunossenescência: envelhecimento do sistema imuneNoch keine Bewertungen
- Apostila de Anatomia e Fisiologia HumanaDokument46 SeitenApostila de Anatomia e Fisiologia HumanaAndreia MonteiroNoch keine Bewertungen
- COP - Extrato de Semente de Uva: Viva Mais e Mais Saudável: Anti-idade para TodosVon EverandCOP - Extrato de Semente de Uva: Viva Mais e Mais Saudável: Anti-idade para TodosNoch keine Bewertungen
- Aula de OligoterapiaDokument12 SeitenAula de OligoterapiaRosa R LimaNoch keine Bewertungen
- GeopatiaDokument2 SeitenGeopatiaridosresende100% (1)
- Educação, Paradoxos e Metamorfoses: Plantas medicinais na escolaVon EverandEducação, Paradoxos e Metamorfoses: Plantas medicinais na escolaBewertung: 5 von 5 Sternen5/5 (2)
- ChlorellaDokument3 SeitenChlorellahelloforwardNoch keine Bewertungen
- Dieta Do Tipo SanguíneoDokument6 SeitenDieta Do Tipo SanguíneoRosa CarvalhoNoch keine Bewertungen
- Quimica Da SaúdeDokument77 SeitenQuimica Da SaúdeMarta SousaNoch keine Bewertungen
- Baço Pancreas RecomendaçõesDokument2 SeitenBaço Pancreas RecomendaçõesPaula Viegas100% (1)
- Anatomia e FisiologiaDokument56 SeitenAnatomia e FisiologiaReginaldo Thuler100% (1)
- Espondilite Anquilosante (Artigo)Dokument11 SeitenEspondilite Anquilosante (Artigo)api-3698831100% (1)
- Sistema ABO e Seu Impacto Na TrasnfDokument15 SeitenSistema ABO e Seu Impacto Na TrasnfEmílio OlímpioNoch keine Bewertungen
- Tratamento de Divertículos Do Tubo DigestivoDokument84 SeitenTratamento de Divertículos Do Tubo DigestivoGiovanna TardinNoch keine Bewertungen
- Cuantica VeterinariaDokument7 SeitenCuantica VeterinariaAnonymous cFCe02Noch keine Bewertungen
- Fisiologia GastrointestinalDokument14 SeitenFisiologia GastrointestinalPaulo Henrique Enzele100% (1)
- Alergias Alimentares: Setores SintomasDokument1 SeiteAlergias Alimentares: Setores SintomasSinara OliverNoch keine Bewertungen
- Compêndio de Produtos Homeopáticos PDFDokument90 SeitenCompêndio de Produtos Homeopáticos PDFrexland1Noch keine Bewertungen
- APENDICITEDokument26 SeitenAPENDICITEÉdina AraújoNoch keine Bewertungen
- ARTIGO A Importância Do Magnésio Na Doença CardiovascularDokument12 SeitenARTIGO A Importância Do Magnésio Na Doença CardiovascularMacielNoch keine Bewertungen
- Energia Protetora - Selo Energético - Loja OficiaDokument1 SeiteEnergia Protetora - Selo Energético - Loja OficiaMarco e Monica C. BNoch keine Bewertungen
- Guia de Uso Encourage - FlowermaxDokument1 SeiteGuia de Uso Encourage - FlowermaxMauroNoch keine Bewertungen
- Medicina Complementar Saiba Quais Tratamentos o SUS Já Oferece - Alex BotsarisDokument5 SeitenMedicina Complementar Saiba Quais Tratamentos o SUS Já Oferece - Alex BotsarisEn100% (1)
- Santo Sarraceno TeaserDokument6 SeitenSanto Sarraceno TeaserErick_Paulino_SouzaNoch keine Bewertungen
- Homeopatia Como Forma de Obter Energia V PDFDokument18 SeitenHomeopatia Como Forma de Obter Energia V PDFAndreaAraujoNoch keine Bewertungen
- Formação IOmet 09-2017Dokument72 SeitenFormação IOmet 09-2017pauloasvNoch keine Bewertungen
- 14 Aula - Fisiologia Do Osso, Cálcio e FósforoDokument4 Seiten14 Aula - Fisiologia Do Osso, Cálcio e Fósforoapi-26429188100% (3)
- O Câncer Tem CuraDokument61 SeitenO Câncer Tem CuraAdriano ValdemirNoch keine Bewertungen
- (Terapias Alternativas) Ciência e Arte É A Iridologia - IrisDiagnoseDokument175 Seiten(Terapias Alternativas) Ciência e Arte É A Iridologia - IrisDiagnoseValéria C Soto100% (1)
- Farmacologia - AnticoncepcionaisDokument20 SeitenFarmacologia - Anticoncepcionaiskahionz100% (2)
- Tudo o Que Voce Precisa Saber Sobre o Metodo SPDokument10 SeitenTudo o Que Voce Precisa Saber Sobre o Metodo SPJosélia Fátima Carnielo DiasNoch keine Bewertungen
- Apostila de IridologiaDokument12 SeitenApostila de IridologiaAna Maria SouzaNoch keine Bewertungen
- Compêndio de Tratamento em CromoterapiaDokument28 SeitenCompêndio de Tratamento em CromoterapiaMircia100% (1)
- Neurotransmissores Diversidade e Funções FormatDokument10 SeitenNeurotransmissores Diversidade e Funções FormatJorge HegelNoch keine Bewertungen
- Drenagem Linfatica Facial e Corporal Uma Revisao de LiteraturaDokument4 SeitenDrenagem Linfatica Facial e Corporal Uma Revisao de LiteraturaMariana Assis DamasioNoch keine Bewertungen
- Discovery Publicações Ervas e Plantas Medicinais 30,08,2021Dokument82 SeitenDiscovery Publicações Ervas e Plantas Medicinais 30,08,2021trato euNoch keine Bewertungen
- SIMPÓSIO HomeopatiaDokument1 SeiteSIMPÓSIO HomeopatiaRuiNoch keine Bewertungen
- 2 - Lectina e LecitinaDokument7 Seiten2 - Lectina e LecitinaJefferson CândidoNoch keine Bewertungen
- Mucunã - Mucuna Pruriens - Natureza BelaDokument8 SeitenMucunã - Mucuna Pruriens - Natureza BelaEduardoCarvalhoNoch keine Bewertungen
- Oligoelementos NaturalmedDokument6 SeitenOligoelementos NaturalmedAntonio de AlexandreNoch keine Bewertungen
- Composição Do Sangue PDFDokument4 SeitenComposição Do Sangue PDFJones CorreaNoch keine Bewertungen
- Sistema Digestivo 2021-22Dokument52 SeitenSistema Digestivo 2021-22depilmed LojaNoch keine Bewertungen
- Neuro - Roteiro de SemiotécnicaDokument14 SeitenNeuro - Roteiro de SemiotécnicaWillian PegoraroNoch keine Bewertungen
- Aveloz e HomeopatDokument7 SeitenAveloz e Homeopatgeeemholo2772Noch keine Bewertungen
- Peroxido de HidroginioDokument2 SeitenPeroxido de Hidroginioajp1000Noch keine Bewertungen
- Eco EnzymaDokument34 SeitenEco EnzymaFrustreidenovoNoch keine Bewertungen
- Um Remédio Incrível para Um Mal Incurável e Outros Males 2Dokument2 SeitenUm Remédio Incrível para Um Mal Incurável e Outros Males 2Geisieli R De Oliveira100% (1)
- Cloreto de MagnesioDokument3 SeitenCloreto de MagnesioadzsrNoch keine Bewertungen
- Peptídeos de Importancia MédicaDokument3 SeitenPeptídeos de Importancia MédicaCarlos AngeloNoch keine Bewertungen
- Como Desintoxicar Seu CorpoDokument16 SeitenComo Desintoxicar Seu CorpoEmerson F LeiteNoch keine Bewertungen
- Ecologia CelularDokument57 SeitenEcologia CelularCarlos Braghini Jr.Noch keine Bewertungen
- Obesidade InfantilDokument2 SeitenObesidade Infantilesf paredão postoNoch keine Bewertungen
- Alcalinização MilagrosaDokument5 SeitenAlcalinização Milagrosaclarindo_gouveiaNoch keine Bewertungen
- Tensão Pre MenstrualDokument13 SeitenTensão Pre MenstrualwesleycaNoch keine Bewertungen
- Anamnese Ortobiomolecular: Ler Antes de PreencherDokument2 SeitenAnamnese Ortobiomolecular: Ler Antes de PreencherEdmar JuniorNoch keine Bewertungen
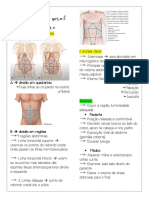
- Exame Fisico AbdominalDokument7 SeitenExame Fisico AbdominalAna Clara GardioliNoch keine Bewertungen
- Fisiologia Do Estresse e Sua InfluenciaDokument13 SeitenFisiologia Do Estresse e Sua InfluenciaEllen TelacioNoch keine Bewertungen
- Alimentação Ancestral e A Dieta Low Carb (Baixo Teor de CarboidratosDokument131 SeitenAlimentação Ancestral e A Dieta Low Carb (Baixo Teor de CarboidratosMarcelasimmerNoch keine Bewertungen
- Exam 694669Dokument8 SeitenExam 694669flavio2015Noch keine Bewertungen
- Prova 1Dokument17 SeitenProva 1Jardel Lemos Thalhofer100% (1)
- Anexo 7 - Documento de Movimentação Do Acervo Documental de 09.10.2009 PDFDokument3 SeitenAnexo 7 - Documento de Movimentação Do Acervo Documental de 09.10.2009 PDFMuseu de História da Medicina do RS - MUHMNoch keine Bewertungen
- Biologia Molecular Básica Estudo DirigidoDokument3 SeitenBiologia Molecular Básica Estudo DirigidoLeonardo GarciaNoch keine Bewertungen
- Kayla Santos - Variação Do MonoibridismoDokument6 SeitenKayla Santos - Variação Do MonoibridismoKayla CpmNoch keine Bewertungen
- Vírus, Pragas e História Presente, Passado e Futuro MICHAEL B A OLDSTONEDokument643 SeitenVírus, Pragas e História Presente, Passado e Futuro MICHAEL B A OLDSTONESergioNoch keine Bewertungen
- FotossinteseDokument19 SeitenFotossinteseMarcos Ricardo LeiteNoch keine Bewertungen
- PCR e RT-QPCR e Suas Aplicações Na Área MédicaDokument10 SeitenPCR e RT-QPCR e Suas Aplicações Na Área MédicaAriadne F.Noch keine Bewertungen
- A Cidos Nucleicos I - Dna e Replicac A oDokument22 SeitenA Cidos Nucleicos I - Dna e Replicac A oVítor S. SilvaNoch keine Bewertungen
- Níveis de OrganizaçãoDokument6 SeitenNíveis de Organizaçãocaua12Noch keine Bewertungen
- Estudo Dirigido 02 - VirologiaDokument3 SeitenEstudo Dirigido 02 - VirologiaAna Silvia Araújo AparícioNoch keine Bewertungen
- A Célula É A Unidade Básica Da VidaDokument8 SeitenA Célula É A Unidade Básica Da VidaAngelica BertolettiNoch keine Bewertungen
- Teste-Módulo 4Dokument5 SeitenTeste-Módulo 4Sara MartinsNoch keine Bewertungen
- GenéticaDokument17 SeitenGenéticaYasminNoch keine Bewertungen
- CEF Texto Informativo - ClonagemDokument4 SeitenCEF Texto Informativo - ClonagemSonia RibeiroNoch keine Bewertungen
- EmbriologiaDokument6 SeitenEmbriologiaLetícia BoniniNoch keine Bewertungen
- MICOBACTÉRIASDokument19 SeitenMICOBACTÉRIASReodino SversutNoch keine Bewertungen
- Livro Embrio 2017Dokument285 SeitenLivro Embrio 2017Gisla Ys100% (1)
- CITOLOGIA SlidesDokument47 SeitenCITOLOGIA SlidesLuciane NevesNoch keine Bewertungen
- PD CitogeneticaDokument4 SeitenPD CitogeneticaLucio MotaNoch keine Bewertungen
- Avaliação II - BiologiaDokument5 SeitenAvaliação II - BiologiaDenis MachadoNoch keine Bewertungen
- Estou Compartilhando o Arquivo 'QUESTOES LUANA' Com VocêDokument17 SeitenEstou Compartilhando o Arquivo 'QUESTOES LUANA' Com Vocêyonara e ana yoeanaNoch keine Bewertungen
- Biogeo11 Teste1Dokument8 SeitenBiogeo11 Teste1clahs rodrigues100% (1)
- Aebg10 B Teste1 PDFDokument2 SeitenAebg10 B Teste1 PDFHelena Alves100% (1)
- Aula 1 - Bioquímica - Medicina UFAC 2018.1 - Fundamentos Da BioquímicaDokument68 SeitenAula 1 - Bioquímica - Medicina UFAC 2018.1 - Fundamentos Da Bioquímica070971weNoch keine Bewertungen
- 1 e 2 Lei de MendelDokument29 Seiten1 e 2 Lei de MendelTulio Paixão de FrançaNoch keine Bewertungen
- Manual Do FacilitadorDokument44 SeitenManual Do FacilitadorSamuel Luís MequeNoch keine Bewertungen
- Voltar Alterar Modo de VisualizaçãoDokument5 SeitenVoltar Alterar Modo de VisualizaçãoJu HobtzNoch keine Bewertungen
- Biologia Atividade Fixação 4Dokument2 SeitenBiologia Atividade Fixação 4Adriana SoaresNoch keine Bewertungen
- Bioinformática 1Dokument12 SeitenBioinformática 1TitosNoch keine Bewertungen
- 21 dias para curar sua vida: Amando a si mesmo trabalhando com o espelhoVon Everand21 dias para curar sua vida: Amando a si mesmo trabalhando com o espelhoBewertung: 4.5 von 5 Sternen4.5/5 (49)
- As 5 linguagens do amor - 3ª edição: Como expressar um compromisso de amor a seu cônjugeVon EverandAs 5 linguagens do amor - 3ª edição: Como expressar um compromisso de amor a seu cônjugeBewertung: 5 von 5 Sternen5/5 (220)
- Fogo no parquinho: Namoro à luz da Palavra de DeusVon EverandFogo no parquinho: Namoro à luz da Palavra de DeusBewertung: 5 von 5 Sternen5/5 (9)
- Deu a louca nos signos: Um guia divertido do zodíaco por AstroloucamenteVon EverandDeu a louca nos signos: Um guia divertido do zodíaco por AstroloucamenteBewertung: 5 von 5 Sternen5/5 (1)
- Psicologia sombria: Poderosas técnicas de controle mental e persuasãoVon EverandPsicologia sombria: Poderosas técnicas de controle mental e persuasãoBewertung: 4 von 5 Sternen4/5 (92)
- Eu te amo, mas não te quero mais: Histórias de fins e recomeçosVon EverandEu te amo, mas não te quero mais: Histórias de fins e recomeçosBewertung: 5 von 5 Sternen5/5 (6)
- Desbloqueie o poder da sua mente: Programe o seu subconsciente para se libertar das dores e inseguranças e transforme a sua vidaVon EverandDesbloqueie o poder da sua mente: Programe o seu subconsciente para se libertar das dores e inseguranças e transforme a sua vidaBewertung: 4.5 von 5 Sternen4.5/5 (226)
- Os Códigos do Mindset da Prosperidade: destrave os bloqueios em sua mente e cresça em todos os aspectos de sua vidaVon EverandOs Códigos do Mindset da Prosperidade: destrave os bloqueios em sua mente e cresça em todos os aspectos de sua vidaNoch keine Bewertungen
- O Verdadeiro Ho'oponopono: Como restaurar sua harmonia, limpar memórias e manifestar milagresVon EverandO Verdadeiro Ho'oponopono: Como restaurar sua harmonia, limpar memórias e manifestar milagresBewertung: 5 von 5 Sternen5/5 (5)
- Focar: Supere a procrastinação e aumente a força de vontade e a atençãoVon EverandFocar: Supere a procrastinação e aumente a força de vontade e a atençãoBewertung: 4.5 von 5 Sternen4.5/5 (53)
- 35 Técnicas e Curiosidades Mentais: Porque a mente também deve evoluirVon Everand35 Técnicas e Curiosidades Mentais: Porque a mente também deve evoluirBewertung: 5 von 5 Sternen5/5 (3)
- Técnicas Proibidas de Manipulação Mental e PersuasãoVon EverandTécnicas Proibidas de Manipulação Mental e PersuasãoBewertung: 5 von 5 Sternen5/5 (3)
- Magic And Gravoboi For Prosperity In 2024 - EnglishVon EverandMagic And Gravoboi For Prosperity In 2024 - EnglishNoch keine Bewertungen
- Almanaque do Pensamento 2024: Guia astrológico para 2024Von EverandAlmanaque do Pensamento 2024: Guia astrológico para 2024Bewertung: 5 von 5 Sternen5/5 (2)
- Descolonizando afetos: Experimentações sobre outras formas de amarVon EverandDescolonizando afetos: Experimentações sobre outras formas de amarBewertung: 5 von 5 Sternen5/5 (6)
- O fim da ansiedade: O segredo bíblico para livrar-se das preocupaçõesVon EverandO fim da ansiedade: O segredo bíblico para livrar-se das preocupaçõesBewertung: 5 von 5 Sternen5/5 (16)
- Crianças desafiadoras: Aprenda como identificar, tratar e contribuir de maneira positiva com crianças que têm Transtorno Opositivo-DesafiadorVon EverandCrianças desafiadoras: Aprenda como identificar, tratar e contribuir de maneira positiva com crianças que têm Transtorno Opositivo-DesafiadorNoch keine Bewertungen
- Descolonizando afetos: Experimentações sobre outras formas de amarVon EverandDescolonizando afetos: Experimentações sobre outras formas de amarBewertung: 5 von 5 Sternen5/5 (1)
- Treinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisVon EverandTreinamento cerebral: Compreendendo inteligência emocional, atenção e muito maisBewertung: 4.5 von 5 Sternen4.5/5 (169)
- Técnicas De Terapia Cognitivo-comportamental (tcc)Von EverandTécnicas De Terapia Cognitivo-comportamental (tcc)Noch keine Bewertungen