Das könnte Ihnen auch gefallen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Vaccines - From Concept To Clinic - L. Paoletti, P. McInnes (CRC, 1999) WW PDFDokument210 SeitenVaccines - From Concept To Clinic - L. Paoletti, P. McInnes (CRC, 1999) WW PDFPatriciaNoch keine Bewertungen
- Janeways Immunobiology 9th EditionDokument19 SeitenJaneways Immunobiology 9th EditionDevin MckayNoch keine Bewertungen
- Greyson 2019Dokument10 SeitenGreyson 2019Ziha Zia Leonita FauziNoch keine Bewertungen
- Home o ProphylaxisDokument26 SeitenHome o ProphylaxisLokir_leika_vllrNoch keine Bewertungen
- Humoral ImmunityDokument86 SeitenHumoral ImmunitySinthiya KanesaratnamNoch keine Bewertungen
- 4life Transfer Factor: Which Product Is Right For Me?Dokument1 Seite4life Transfer Factor: Which Product Is Right For Me?jhimsyNoch keine Bewertungen
- Passive Voice PDFDokument2 SeitenPassive Voice PDFalya nafaNoch keine Bewertungen
- Koch's PostulatesDokument5 SeitenKoch's PostulatessaaisNoch keine Bewertungen
- Archer (Staphylococcus Aureus)Dokument3 SeitenArcher (Staphylococcus Aureus)_meiantiNoch keine Bewertungen
- Campbell Smith - Harris032311Dokument2 SeitenCampbell Smith - Harris032311Heather X RhodesNoch keine Bewertungen
- Why GILD Remdesivir Is The Only Relevant Coronavirus DrugDokument8 SeitenWhy GILD Remdesivir Is The Only Relevant Coronavirus Drugjulia skripka-serry100% (2)
- REVIEW OF MEDICAL MICROBIOLOGY-Twelfth Edition PDFDokument2 SeitenREVIEW OF MEDICAL MICROBIOLOGY-Twelfth Edition PDFSai Venkata Suresh VarmaNoch keine Bewertungen
- Guide To Biologic Therapy 1Dokument24 SeitenGuide To Biologic Therapy 1Andra GarbeaNoch keine Bewertungen
- Atopic Dermatitis - The Epidemiology, Causes and Prevention of Atopic Eczema (PDFDrive)Dokument289 SeitenAtopic Dermatitis - The Epidemiology, Causes and Prevention of Atopic Eczema (PDFDrive)NguyenDinh NinhNoch keine Bewertungen
- Dengue Final Case Report Ready - OdtDokument5 SeitenDengue Final Case Report Ready - Odtroshan bhandariNoch keine Bewertungen
- F. KG - Prosiding - Sri Hernawati - Penatalaksanaan Fisured Tongue Disertai DentureDokument8 SeitenF. KG - Prosiding - Sri Hernawati - Penatalaksanaan Fisured Tongue Disertai DentureAlfan MaulanaNoch keine Bewertungen
- Rife Consolidated Annotated Frequency List - EMR Labs, LLCDokument19 SeitenRife Consolidated Annotated Frequency List - EMR Labs, LLCMatimba SimangoNoch keine Bewertungen
- Symptoms of AsthmaDokument4 SeitenSymptoms of AsthmaAthina Maricar CabaseNoch keine Bewertungen
- JURNALDokument9 SeitenJURNALagus nugrahaNoch keine Bewertungen
- Student Medical RecordDokument3 SeitenStudent Medical Recordhamsa RasbiehNoch keine Bewertungen
- Krok 1 - 2015 (Microbiology)Dokument17 SeitenKrok 1 - 2015 (Microbiology)mayna ynaNoch keine Bewertungen
- Meningococcal Disease: What College Communities Need To KnowDokument2 SeitenMeningococcal Disease: What College Communities Need To KnowSinclair Broadcast Group - EugeneNoch keine Bewertungen
- Tan, Jolwin Laugo 2361052561Dokument6 SeitenTan, Jolwin Laugo 2361052561JolwinTanNoch keine Bewertungen
- Immunology & Immunotechnology - 1Dokument40 SeitenImmunology & Immunotechnology - 1jihovajayraNoch keine Bewertungen
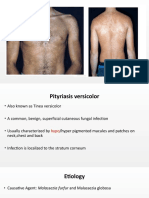
- Pityriasis VersicolorDokument21 SeitenPityriasis VersicolorRahul SrivastavaNoch keine Bewertungen
- Micro 5Dokument51 SeitenMicro 5NeviNoch keine Bewertungen
- Test Report:: Patient Name:Mr Bahu Anil Kumar Sahu Patient IDDokument1 SeiteTest Report:: Patient Name:Mr Bahu Anil Kumar Sahu Patient IDANILNoch keine Bewertungen
- Dengue Hemorrhagic Fever PathophysiologyDokument4 SeitenDengue Hemorrhagic Fever PathophysiologyKirk Espanol BigstoneNoch keine Bewertungen
- Clinical Presentation On ADDDokument39 SeitenClinical Presentation On ADDSREEDEVI T SURESHNoch keine Bewertungen
- Yashika RajDokument1 SeiteYashika RajNitin RajeevNoch keine Bewertungen