Das könnte Ihnen auch gefallen
- In Silico Research in Drug DiscoveryDokument4 SeitenIn Silico Research in Drug Discoverybelgu amineNoch keine Bewertungen
- Transgenic ValidationDokument22 SeitenTransgenic ValidationSyama J.S33% (6)
- Presented By: Anand Sagar Tiwari M.Pharm (Second Semester)Dokument56 SeitenPresented By: Anand Sagar Tiwari M.Pharm (Second Semester)Dawit BirhanuNoch keine Bewertungen
- Application of Biotechnology and Engineering in Veterinary Science StudentsDokument8 SeitenApplication of Biotechnology and Engineering in Veterinary Science StudentsAdnan AhmedNoch keine Bewertungen
- 1 s2.0 S0753332221006156 MainDokument18 Seiten1 s2.0 S0753332221006156 MainBen DresimNoch keine Bewertungen
- PH DD Lecture ReviewerDokument4 SeitenPH DD Lecture ReviewerJoenly CardenasNoch keine Bewertungen
- Early Drug Discovery ProcessDokument58 SeitenEarly Drug Discovery ProcessFrietzyl Mae Generalao100% (1)
- Molecular Methods Used in Clinical Laboratory: Prospects and PitfallsDokument8 SeitenMolecular Methods Used in Clinical Laboratory: Prospects and PitfallsRon VitorNoch keine Bewertungen
- Ni Hms 612555Dokument14 SeitenNi Hms 612555romanNoch keine Bewertungen
- Target Identification and Mechanism of Action in Chemical Biology and Drug DiscoveryDokument9 SeitenTarget Identification and Mechanism of Action in Chemical Biology and Drug DiscoveryPietro DelreNoch keine Bewertungen
- Antibody GenerationDokument7 SeitenAntibody Generationapi-3811432Noch keine Bewertungen
- Reverse PharmacolgoyDokument55 SeitenReverse Pharmacolgoyعبدالله الجزارNoch keine Bewertungen
- Biomarker - Protein Biomarker Discovery and Validation - The Long and Uncertain Path To Clinical UtilityDokument13 SeitenBiomarker - Protein Biomarker Discovery and Validation - The Long and Uncertain Path To Clinical UtilityCher IshNoch keine Bewertungen
- 4bpharm FindDokument20 Seiten4bpharm FindA.R.N.U TECH CHANNELNoch keine Bewertungen
- Cell PaintingDokument18 SeitenCell PaintingdupuytrenNoch keine Bewertungen
- Lindsay Nat Rev DD 2003Dokument8 SeitenLindsay Nat Rev DD 2003douniahamouda87Noch keine Bewertungen
- Cellular Biomechanics IIDokument14 SeitenCellular Biomechanics IILeticiaNoch keine Bewertungen
- New Approaches To Molecular Cancer TherapyDokument12 SeitenNew Approaches To Molecular Cancer TherapyGourav DasNoch keine Bewertungen
- Precision Metagenomics Ebook - M GL 01787Dokument26 SeitenPrecision Metagenomics Ebook - M GL 01787aniioanaNoch keine Bewertungen
- The Necessity of and Strategies For Improving Confidence in The Accuracy of Western BlotsDokument23 SeitenThe Necessity of and Strategies For Improving Confidence in The Accuracy of Western BlotsDouglas SantosNoch keine Bewertungen
- Kingsmore - 2006 - Multiplexed Protein Measurement Technologies andDokument11 SeitenKingsmore - 2006 - Multiplexed Protein Measurement Technologies andLaura GarciaNoch keine Bewertungen
- Biomarkers of Infection and SepsisDokument12 SeitenBiomarkers of Infection and SepsisnuelitopNoch keine Bewertungen
- High-Throughput Approaches To Uncover SynergisticDokument12 SeitenHigh-Throughput Approaches To Uncover SynergisticRodrigo MaxNoch keine Bewertungen
- TS Scripps Lab Recombinant Proteins Ebook D6Dokument8 SeitenTS Scripps Lab Recombinant Proteins Ebook D6dominiqueNoch keine Bewertungen
- 4.2 Drug Development Series - Preclinical - Pharmacology-1-1Dokument20 Seiten4.2 Drug Development Series - Preclinical - Pharmacology-1-1Rhythm BhardwajNoch keine Bewertungen
- Target Validation in Drug DiscoveryVon EverandTarget Validation in Drug DiscoveryBrian W. MetcalfNoch keine Bewertungen
- Benoit 2018Dokument15 SeitenBenoit 2018Rohan PandeyNoch keine Bewertungen
- Antibody Validation For Protein Expression On Tissue Slides - A Protocol For Immunohistochemistry PDFDokument10 SeitenAntibody Validation For Protein Expression On Tissue Slides - A Protocol For Immunohistochemistry PDFOscar Alonso LunaNoch keine Bewertungen
- ELISA in The Multiplex Era Potential and PitfallsDokument17 SeitenELISA in The Multiplex Era Potential and PitfallsHameed SabahNoch keine Bewertungen
- Identification of Novel p53 Pathway Activating Small-Molecule Compounds Reveals Unexpected Similarities With Known Therapeutic AgentsDokument11 SeitenIdentification of Novel p53 Pathway Activating Small-Molecule Compounds Reveals Unexpected Similarities With Known Therapeutic Agentshttps://nootropicsjet.com/Noch keine Bewertungen
- Binding Kinetics in Drug discovery-BMG LabDokument25 SeitenBinding Kinetics in Drug discovery-BMG LabRichard GabrielNoch keine Bewertungen
- Drug Development: FDA's Definition of A New DrugDokument11 SeitenDrug Development: FDA's Definition of A New DrugSophie MendezNoch keine Bewertungen
- Review Article Introduction To Chemical Proteomics For Drug Discovery and DevelopmentDokument9 SeitenReview Article Introduction To Chemical Proteomics For Drug Discovery and DevelopmentPaviliuc RalucaNoch keine Bewertungen
- Highly Multiplexed Immunofluorescence Imaging of Human Tissues and Tumors Using T-CyCIF and Conventional Optical MicroscopesDokument46 SeitenHighly Multiplexed Immunofluorescence Imaging of Human Tissues and Tumors Using T-CyCIF and Conventional Optical MicroscopesCavin PengNoch keine Bewertungen
- Identifying and Quantifying Heterogeneity in High Content Analysis: Application of Heterogeneity Indices To Drug DiscoveryDokument16 SeitenIdentifying and Quantifying Heterogeneity in High Content Analysis: Application of Heterogeneity Indices To Drug DiscoveryMuhammad SatrioNoch keine Bewertungen
- Current and Potential Immunohistochemical Biomarkers For P 2021 Seminars inDokument9 SeitenCurrent and Potential Immunohistochemical Biomarkers For P 2021 Seminars inClaudinete SouzaNoch keine Bewertungen
- Proteomics in Drug Discovery: Jack H. Wang and Rodney M. HewickDokument5 SeitenProteomics in Drug Discovery: Jack H. Wang and Rodney M. HewickBe EducatedNoch keine Bewertungen
- Pi Is 2213219820312435Dokument3 SeitenPi Is 2213219820312435BrilliantNoch keine Bewertungen
- Topic 2 - Drug Discovery and DesignDokument53 SeitenTopic 2 - Drug Discovery and DesignSenaima Marie Hilario BantosaNoch keine Bewertungen
- 1 s2.0 S2452231718300034 MainDokument6 Seiten1 s2.0 S2452231718300034 MainRofiqi RofiqNoch keine Bewertungen
- Current Trends in Drug Discovery Target Identifica PDFDokument5 SeitenCurrent Trends in Drug Discovery Target Identifica PDFKashish GuptaNoch keine Bewertungen
- Identificación y Validación de Objetivos Farmacológicos Controlados GenéticamenteDokument13 SeitenIdentificación y Validación de Objetivos Farmacológicos Controlados GenéticamenteBrayanTrujilloNoch keine Bewertungen
- A Genotype-Phenotype Research Strategy For SchizophreniaDokument13 SeitenA Genotype-Phenotype Research Strategy For Schizophrenianitika lalNoch keine Bewertungen
- Editorials: (4), While Inducing A Longer-Lived Protein (Insulin-LikeDokument2 SeitenEditorials: (4), While Inducing A Longer-Lived Protein (Insulin-LikeAriSuandiNoch keine Bewertungen
- Principle and Techniques of Immunohistochemistry A Review PDFDokument7 SeitenPrinciple and Techniques of Immunohistochemistry A Review PDFMohammed R.HusseinNoch keine Bewertungen
- Combination Drug Testing DeviceDokument24 SeitenCombination Drug Testing DeviceTaha AzadNoch keine Bewertungen
- Mol Methods To Antibacterial ResisDokument18 SeitenMol Methods To Antibacterial ResisPisces EdgeNoch keine Bewertungen
- Biomedicine & Pharmacotherapy: Mohammad Yousef Memar, Hossein Bannazadeh Baghi TDokument8 SeitenBiomedicine & Pharmacotherapy: Mohammad Yousef Memar, Hossein Bannazadeh Baghi TNurul Huda KowitaNoch keine Bewertungen
- Measuring Cancer Drug Sensitivity and Resistance in Cultured Cells: Cancer Drug Sensitivity and Resistance in CellsDokument21 SeitenMeasuring Cancer Drug Sensitivity and Resistance in Cultured Cells: Cancer Drug Sensitivity and Resistance in CellsFarhana AnuarNoch keine Bewertungen
- HHS Public Access: Drug Target Identification in Protozoan ParasitesDokument21 SeitenHHS Public Access: Drug Target Identification in Protozoan ParasitesJKayckeNoch keine Bewertungen
- Principles of Early Drug DiscoveryDokument11 SeitenPrinciples of Early Drug DiscoveryAmrit Karmarkar RPhNoch keine Bewertungen
- CTI2 9 E1183Dokument26 SeitenCTI2 9 E1183PabloNoch keine Bewertungen
- 2012 ESMO Handbook of Clinical Pharmacology of Anti Cancer Agents Chapter 1Dokument8 Seiten2012 ESMO Handbook of Clinical Pharmacology of Anti Cancer Agents Chapter 1Md Yusuf AnsariNoch keine Bewertungen
- Phenotypic Screens As A Renewed Approach For Drug Discovery: Wei Zheng, Natasha Thorne and John C. MckewDokument7 SeitenPhenotypic Screens As A Renewed Approach For Drug Discovery: Wei Zheng, Natasha Thorne and John C. MckewArchie CabacheteNoch keine Bewertungen
- BMMDokument19 SeitenBMMRitika agrawalNoch keine Bewertungen
- Chapter 5 - Protein TherapeuticsDokument68 SeitenChapter 5 - Protein TherapeuticsNew MusicNoch keine Bewertungen
- Drug Development - Credit 2Dokument69 SeitenDrug Development - Credit 2shubham sabaleNoch keine Bewertungen
- Implementation of Proteomics in Clinical TrialsDokument12 SeitenImplementation of Proteomics in Clinical TrialsFarmaceutico RaulNoch keine Bewertungen
- 1 s2.0 S1387265607130040 MainDokument30 Seiten1 s2.0 S1387265607130040 Mainsecret walang clueNoch keine Bewertungen
- Pharmacology in Drug Discovery: Understanding Drug ResponseVon EverandPharmacology in Drug Discovery: Understanding Drug ResponseNoch keine Bewertungen
- Coarse Dispersions: By: Daisy Marie A. Tecson and Claire R. LagoDokument28 SeitenCoarse Dispersions: By: Daisy Marie A. Tecson and Claire R. LagoArchie CabacheteNoch keine Bewertungen
- Chapter 15 - Anticholinergic and Neuroleptic DrugsDokument1 SeiteChapter 15 - Anticholinergic and Neuroleptic DrugsArchie CabacheteNoch keine Bewertungen
- RHEOLOGYDokument18 SeitenRHEOLOGYArchie CabacheteNoch keine Bewertungen
- Solubility and Distribution PhenomenaDokument28 SeitenSolubility and Distribution PhenomenaArchie Cabachete100% (1)
- PhypharmDokument23 SeitenPhypharmArchie CabacheteNoch keine Bewertungen
- 2018 Value-in-HeDokument7 Seiten2018 Value-in-HeArchie CabacheteNoch keine Bewertungen
- CHOLERADokument5 SeitenCHOLERAArchie CabacheteNoch keine Bewertungen
- Cosmetic Remove Hair Skin HumanDokument9 SeitenCosmetic Remove Hair Skin HumanArchie CabacheteNoch keine Bewertungen
- Hospital Report by P.LDokument16 SeitenHospital Report by P.LArchie CabacheteNoch keine Bewertungen
- Why Is There A Need of Dosage FormDokument2 SeitenWhy Is There A Need of Dosage FormArchie CabacheteNoch keine Bewertungen
- Environmental Pollution and Waste: Air, Water, and Land MediaDokument32 SeitenEnvironmental Pollution and Waste: Air, Water, and Land MediaArchie CabacheteNoch keine Bewertungen
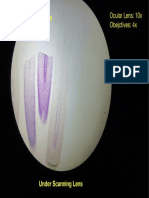
- Allium Cepa Root Tip L.S: Ocular Lens: 10x Obejctives: 4xDokument1 SeiteAllium Cepa Root Tip L.S: Ocular Lens: 10x Obejctives: 4xArchie CabacheteNoch keine Bewertungen
- Code of Ethics For PharmacistsDokument1 SeiteCode of Ethics For PharmacistsArchie CabacheteNoch keine Bewertungen
- Part I: Anatomy and Physiology of The Digestive SytemDokument8 SeitenPart I: Anatomy and Physiology of The Digestive SytemArchie CabacheteNoch keine Bewertungen
- The Role of Planning As A Fundamental Management Function For Achieving Effectiveness in Business OrganizationsDokument10 SeitenThe Role of Planning As A Fundamental Management Function For Achieving Effectiveness in Business OrganizationsArchie CabacheteNoch keine Bewertungen
- The Four TemperamentsDokument30 SeitenThe Four TemperamentsArchie CabacheteNoch keine Bewertungen
- Classifications of Bones: Gross AnatomyDokument6 SeitenClassifications of Bones: Gross AnatomyArchie CabacheteNoch keine Bewertungen
- Pharmacist's Cheat Sheet For Obese Patients: I. ObesityDokument6 SeitenPharmacist's Cheat Sheet For Obese Patients: I. ObesityArchie CabacheteNoch keine Bewertungen
- Objectives:: Exercise PersonDokument11 SeitenObjectives:: Exercise PersonArchie CabacheteNoch keine Bewertungen
- European Journal of Integrative Medicine: Kylie O 'Brien TDokument7 SeitenEuropean Journal of Integrative Medicine: Kylie O 'Brien TArchie CabacheteNoch keine Bewertungen
- Pharmacy and Therapeutics Committee Policies and ProceduresDokument21 SeitenPharmacy and Therapeutics Committee Policies and ProceduresArchie CabacheteNoch keine Bewertungen
- Journal of The American Pharmacists Association: Advances in Pharmacy PracticeDokument9 SeitenJournal of The American Pharmacists Association: Advances in Pharmacy PracticeArchie CabacheteNoch keine Bewertungen
- Using CREATE For AssignmentsDokument2 SeitenUsing CREATE For AssignmentsArchie CabacheteNoch keine Bewertungen
- Journal of King Saud University - ScienceDokument6 SeitenJournal of King Saud University - ScienceArchie CabacheteNoch keine Bewertungen
- Laboratory Name Location: Physics Department Laboratory Policies Page 1 of 9Dokument9 SeitenLaboratory Name Location: Physics Department Laboratory Policies Page 1 of 9Archie CabacheteNoch keine Bewertungen
- Phenotypic Screens As A Renewed Approach For Drug Discovery: Wei Zheng, Natasha Thorne and John C. MckewDokument7 SeitenPhenotypic Screens As A Renewed Approach For Drug Discovery: Wei Zheng, Natasha Thorne and John C. MckewArchie CabacheteNoch keine Bewertungen
- 10 - Preparation of Reagents and FormulasDokument3 Seiten10 - Preparation of Reagents and FormulasArchie CabacheteNoch keine Bewertungen
- Laboratory-Procedure-Policies-Guidelines 2Dokument7 SeitenLaboratory-Procedure-Policies-Guidelines 2Archie CabacheteNoch keine Bewertungen
- Malignant Neoplasm (Ovarian Cancer)Dokument4 SeitenMalignant Neoplasm (Ovarian Cancer)nursing concept mapsNoch keine Bewertungen
- Heat Sterilisation 1Dokument27 SeitenHeat Sterilisation 1romiifreeNoch keine Bewertungen
- Organisms and Environment - IDokument52 SeitenOrganisms and Environment - IFatima AnsariNoch keine Bewertungen
- Primary Motor Cortex Changes After Amputation Correlate With Phantom Limb Pain and The Ability To Move The Phantom Limb 2016 NeuroImageDokument11 SeitenPrimary Motor Cortex Changes After Amputation Correlate With Phantom Limb Pain and The Ability To Move The Phantom Limb 2016 NeuroImageLuís EduardoNoch keine Bewertungen
- Biodanza by Rolando Toro in EnglishDokument90 SeitenBiodanza by Rolando Toro in EnglishJohn Jamieson100% (3)
- Noel Et Al 2016Dokument17 SeitenNoel Et Al 2016Cass CainNoch keine Bewertungen
- 4.1 Geological Time Periods Study MaterialDokument12 Seiten4.1 Geological Time Periods Study MaterialGadgetGlitchKillNoch keine Bewertungen
- Example of Declaration of Biological Shipments FormDokument2 SeitenExample of Declaration of Biological Shipments Formrubana reazNoch keine Bewertungen
- Bang-Bang and Singular in BiologyDokument45 SeitenBang-Bang and Singular in Biologyva3ttnNoch keine Bewertungen
- A Radiologist's Guide To The 2021 WHO Central Nervous System Tumor ClassificationDokument15 SeitenA Radiologist's Guide To The 2021 WHO Central Nervous System Tumor ClassificationMANGNoch keine Bewertungen
- CHEMOSTATDokument2 SeitenCHEMOSTATSamantha SwiftNoch keine Bewertungen
- Calluna Vulgaris UsesDokument1 SeiteCalluna Vulgaris UsesBekaNoch keine Bewertungen
- PhysioEx Exercise 9 Activity 2Dokument3 SeitenPhysioEx Exercise 9 Activity 2PatriNoch keine Bewertungen
- ML Diabetes IeeeDokument12 SeitenML Diabetes IeeeArry A100% (1)
- Chapter 6: Cell Division: 6.3: MEIOSISDokument29 SeitenChapter 6: Cell Division: 6.3: MEIOSISMohammed RidzuwanNoch keine Bewertungen
- Cell Disruption MethodsDokument19 SeitenCell Disruption MethodsMalek Marry AnneNoch keine Bewertungen
- Armando Alducin - Creacion o EvolucionDokument111 SeitenArmando Alducin - Creacion o EvolucionmarianNoch keine Bewertungen
- 9700 - MW - Script F - P3Dokument20 Seiten9700 - MW - Script F - P3Y. PurwandariNoch keine Bewertungen
- PS ReviewerDokument3 SeitenPS ReviewerIt’s yanaNoch keine Bewertungen
- 21 - 6182 - e (C) - PF1 (M) - F (P) - PF1 (PP) - Pfa (PR)Dokument3 Seiten21 - 6182 - e (C) - PF1 (M) - F (P) - PF1 (PP) - Pfa (PR)Vinna KusumawatiNoch keine Bewertungen
- Listado de Especies Maldi Biotyper - Enero 2014Dokument52 SeitenListado de Especies Maldi Biotyper - Enero 2014Elias ENoch keine Bewertungen
- Plants Used To Treat Various Mental Illnesses in Ethiopia A Systematic ReviewDokument26 SeitenPlants Used To Treat Various Mental Illnesses in Ethiopia A Systematic ReviewSIYUM ABAYINEHNoch keine Bewertungen
- Daftar Pustaka 12 September 2016Dokument9 SeitenDaftar Pustaka 12 September 2016Irfan AchmadNoch keine Bewertungen
- Troubleshooting V1.1 11 07 2016 ENGDokument24 SeitenTroubleshooting V1.1 11 07 2016 ENGWilfredo RodriguezNoch keine Bewertungen
- Environmental Monitoring Report-Oktober 2016Dokument31 SeitenEnvironmental Monitoring Report-Oktober 2016azuraidaNoch keine Bewertungen
- Radiation Safety Manual (Duke Univ Laboratory, 2001)Dokument54 SeitenRadiation Safety Manual (Duke Univ Laboratory, 2001)AndréRochaNoch keine Bewertungen
- Gensoc Midterm Theories of Gender Identity DevelopmentDokument5 SeitenGensoc Midterm Theories of Gender Identity DevelopmentDevon DebarrasNoch keine Bewertungen
- WEEK 16 LABORATORY EXERCISE Special SensesDokument7 SeitenWEEK 16 LABORATORY EXERCISE Special SensesGelo AlonzoNoch keine Bewertungen
- 16 Isolation of Chlorophyll and CarotenoidDokument4 Seiten16 Isolation of Chlorophyll and CarotenoidthissisnateNoch keine Bewertungen
- 3 高溫高壓蒸氣滅菌器之介紹、選用與檢測Dokument39 Seiten3 高溫高壓蒸氣滅菌器之介紹、選用與檢測郭哲宏Noch keine Bewertungen