Das könnte Ihnen auch gefallen
- %recognition Accuracy: Speech Recognition Is The Translation of Spoken Words Into TextDokument8 Seiten%recognition Accuracy: Speech Recognition Is The Translation of Spoken Words Into TextPradeep KumarNoch keine Bewertungen
- AbstDokument2 SeitenAbstPradeep KumarNoch keine Bewertungen
- 1) Program For Blinking Leds Connected To Ports P2 and P3Dokument5 Seiten1) Program For Blinking Leds Connected To Ports P2 and P3Pradeep KumarNoch keine Bewertungen
- L9 8upDokument7 SeitenL9 8upPradeep KumarNoch keine Bewertungen
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Eggs Fase 1Dokument3 SeitenEggs Fase 1Gustav MolMedNoch keine Bewertungen
- Immunoproteasome: Dania Mohammad Abu NawasDokument27 SeitenImmunoproteasome: Dania Mohammad Abu NawasDania MohNoch keine Bewertungen
- US Special Virus Program Progress Report 8 1971 PDFDokument378 SeitenUS Special Virus Program Progress Report 8 1971 PDFst_spyridonNoch keine Bewertungen
- Origin of Eukaryotic Cell PDFDokument8 SeitenOrigin of Eukaryotic Cell PDFaco_golarNoch keine Bewertungen
- Is Childhood A Universal Experience?Dokument8 SeitenIs Childhood A Universal Experience?camilo jamesNoch keine Bewertungen
- Biochemistry of GlycoproteinDokument6 SeitenBiochemistry of GlycoproteinMahathir Mohmed100% (7)
- Substance Id enDokument118 SeitenSubstance Id ensmaps001Noch keine Bewertungen
- Intraspecific CompetitionDokument2 SeitenIntraspecific CompetitionZainab MahmudNoch keine Bewertungen
- Timeline of Appearance of Life FormsDokument4 SeitenTimeline of Appearance of Life FormsMaria Andrea CosicoNoch keine Bewertungen
- Lazcano y Pereto. 2021. BioSystemsDokument6 SeitenLazcano y Pereto. 2021. BioSystemsRenato AmosNoch keine Bewertungen
- Gagne Information Processing TheoryDokument36 SeitenGagne Information Processing TheoryHidayatul MabrurNoch keine Bewertungen
- Postdoc Cover Letter SampleDokument7 SeitenPostdoc Cover Letter Samplevofysyv1z1v3100% (1)
- Mob Ot 31753002739420Dokument276 SeitenMob Ot 31753002739420fabriziozaraNoch keine Bewertungen
- Growth Kinetics 2014Dokument33 SeitenGrowth Kinetics 2014Ivan Aulia BigbandNoch keine Bewertungen
- BIO MCQS by IlmiDokument67 SeitenBIO MCQS by IlmiZee waqarNoch keine Bewertungen
- Semi-Detailed Lesson Plan For Cellular RespirationDokument2 SeitenSemi-Detailed Lesson Plan For Cellular RespirationZaifel Pacillos100% (5)
- PROTOCHORDATESDokument4 SeitenPROTOCHORDATESapi-26570979100% (1)
- Duchenne Muscular DystrophyDokument32 SeitenDuchenne Muscular DystrophyMr.P.Ramesh, Faculty of Physiotherapy, SRUNoch keine Bewertungen
- BOOK Embodiment in Psychotherapy - A Practitioner's Guide-Springer International Publishing (2018)Dokument386 SeitenBOOK Embodiment in Psychotherapy - A Practitioner's Guide-Springer International Publishing (2018)PAOLA148Noch keine Bewertungen
- 1,4 ButanediolDokument6 Seiten1,4 ButanediolVictor VikeneNoch keine Bewertungen
- Bye Bye Bison PART I Ed 16Dokument4 SeitenBye Bye Bison PART I Ed 16dquythinh6211Noch keine Bewertungen
- Bio 1M03Dokument73 SeitenBio 1M03brady47Noch keine Bewertungen
- 9780702072987-Book ChapterDokument2 Seiten9780702072987-Book ChaptervisiniNoch keine Bewertungen
- Ecozone Dichotomous KeyDokument3 SeitenEcozone Dichotomous Keyqmtkqz72vhNoch keine Bewertungen
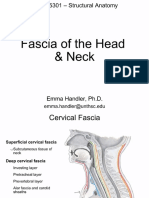
- Fascia of The Head & NeckDokument8 SeitenFascia of The Head & NeckMahesh Jung KhadkaNoch keine Bewertungen
- Comparative Analysis of Agrobiological Traits of Durum (T. Durum Desf.) and Bread Wheat (T. Aestivum L.) Varieties in The Karabakh RegionDokument9 SeitenComparative Analysis of Agrobiological Traits of Durum (T. Durum Desf.) and Bread Wheat (T. Aestivum L.) Varieties in The Karabakh RegionPublisher NasirNoch keine Bewertungen
- Eysenck Personality TheoryDokument11 SeitenEysenck Personality TheoryKarthika RsNoch keine Bewertungen
- Hongos Del ZamoranoDokument6 SeitenHongos Del ZamoranoAnahí GallegosNoch keine Bewertungen
- Eucalyptol 1Dokument10 SeitenEucalyptol 1NadiyahNoch keine Bewertungen
- Two Week Learning Plan ScienceDokument4 SeitenTwo Week Learning Plan ScienceMaria Ramona FelixNoch keine Bewertungen