Das könnte Ihnen auch gefallen
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Natures Hidden Health MiracleDokument38 SeitenNatures Hidden Health MiracleGorneau100% (2)
- Nervous System NotesDokument6 SeitenNervous System NotesAlex Whitwam100% (3)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Recognizing and Treating Breathing DisordersDokument302 SeitenRecognizing and Treating Breathing DisordersJun Kim100% (2)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- PathologyDokument36 SeitenPathologySayeed KhanNoch keine Bewertungen
- Diagnosis For Complete DentureDokument33 SeitenDiagnosis For Complete Dentureyabhatia100% (3)
- Movement DisordersDokument10 SeitenMovement DisordersAli 10Noch keine Bewertungen
- ImageJ Manual For Image Processing and Analysis.Dokument45 SeitenImageJ Manual For Image Processing and Analysis.acr3656100% (1)
- Do The Activity or Answer The Questions Below in Your NotebookDokument3 SeitenDo The Activity or Answer The Questions Below in Your NotebookBern Patrick BautistaNoch keine Bewertungen
- Zinc and Immune Function PDFDokument17 SeitenZinc and Immune Function PDF300r100% (1)
- Cortical Dipole and Clinical SignificanceDokument15 SeitenCortical Dipole and Clinical SignificanceManamitaNoch keine Bewertungen
- Ultrathin Cryosectioning Immunogold Labelling: A Practical Introduction. Manual CMC Course Utrecht 2012Dokument83 SeitenUltrathin Cryosectioning Immunogold Labelling: A Practical Introduction. Manual CMC Course Utrecht 2012acr3656Noch keine Bewertungen
- Basic Photoshop For Electron Microscopy 2014 by Eric Miller Microscopy & Imaging Specialist at Northwestern UniversityDokument91 SeitenBasic Photoshop For Electron Microscopy 2014 by Eric Miller Microscopy & Imaging Specialist at Northwestern Universityacr3656Noch keine Bewertungen
- ImmunoGold. Applications and Techniques2012 EDITIONDokument46 SeitenImmunoGold. Applications and Techniques2012 EDITIONacr3656Noch keine Bewertungen
- The Use of The IonizerDokument1 SeiteThe Use of The Ionizeracr3656Noch keine Bewertungen
- Abcam Protocols Book 2010Dokument82 SeitenAbcam Protocols Book 2010acr3656Noch keine Bewertungen
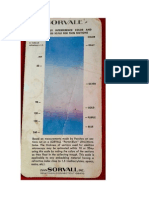
- Sorvall Continuous Interference Colour and Thickness Scale For Thin SectionsDokument1 SeiteSorvall Continuous Interference Colour and Thickness Scale For Thin Sectionsacr3656Noch keine Bewertungen
- Prerequisites For Immunoelectron MicrosDokument3 SeitenPrerequisites For Immunoelectron Microsacr3656Noch keine Bewertungen
- High Pressure Freezing of Rust Infected Plant LeavesDokument12 SeitenHigh Pressure Freezing of Rust Infected Plant Leavesacr3656Noch keine Bewertungen
- Seeing Is Believing: Use of Antibodies in Immunocytochemistry and in Situ HybridizationDokument17 SeitenSeeing Is Believing: Use of Antibodies in Immunocytochemistry and in Situ Hybridizationacr3656Noch keine Bewertungen
- Combined Deparaffinization and Target RetrievalDokument2 SeitenCombined Deparaffinization and Target Retrievalacr3656Noch keine Bewertungen
- Coated GridsDokument2 SeitenCoated Gridsacr3656Noch keine Bewertungen
- Optimization of Immunohistochemical ReactionsDokument6 SeitenOptimization of Immunohistochemical Reactionsacr3656Noch keine Bewertungen
- Combined Use of Confocal Laser Scanning Microscopy and Transmission ElectronDokument9 SeitenCombined Use of Confocal Laser Scanning Microscopy and Transmission Electronacr3656Noch keine Bewertungen
- Proteolytic Pre-Treatment Antigen RetrievalDokument1 SeiteProteolytic Pre-Treatment Antigen Retrievalacr3656Noch keine Bewertungen
- IHC Protocol GuideDokument116 SeitenIHC Protocol Guideacr3656Noch keine Bewertungen
- Combined Proteolytic Pre-Treatment and HIERDokument2 SeitenCombined Proteolytic Pre-Treatment and HIERacr3656Noch keine Bewertungen
- Water Bath Antigen RetrievalDokument3 SeitenWater Bath Antigen Retrievalacr3656Noch keine Bewertungen
- Microwave Oven Antigen RetrievalDokument2 SeitenMicrowave Oven Antigen Retrievalacr3656Noch keine Bewertungen
- Antibody Validation - A Review.Dokument8 SeitenAntibody Validation - A Review.acr3656Noch keine Bewertungen
- Pressure Cooking Antigen RetrievalDokument1 SeitePressure Cooking Antigen Retrievalacr3656Noch keine Bewertungen
- Constructing Tissue Micro Arrays Without PrefabricatingDokument5 SeitenConstructing Tissue Micro Arrays Without Prefabricatingacr3656Noch keine Bewertungen
- Guidelines For Appropriate Controls For ImmunohistochemistryDokument3 SeitenGuidelines For Appropriate Controls For Immunohistochemistryacr3656Noch keine Bewertungen
- A Fixed, Paraffin-Processed Cell Line StandardDokument9 SeitenA Fixed, Paraffin-Processed Cell Line Standardacr3656Noch keine Bewertungen
- Control of AutoFluorescence of Archival FixedDokument7 SeitenControl of AutoFluorescence of Archival Fixedacr3656Noch keine Bewertungen
- Fine Structure ImmunocytochemistryDokument242 SeitenFine Structure Immunocytochemistryacr3656Noch keine Bewertungen
- Ultrathin Cryo-Sectioning and Immuno - Gold LabelingDokument46 SeitenUltrathin Cryo-Sectioning and Immuno - Gold Labelingacr3656100% (1)
- Live-Cell Microscopy - Tips and ToolsDokument15 SeitenLive-Cell Microscopy - Tips and Toolsacr3656Noch keine Bewertungen
- Choosing A Secondary Antibody: A Guide To Fragment SpecificityDokument4 SeitenChoosing A Secondary Antibody: A Guide To Fragment Specificityacr3656Noch keine Bewertungen
- Technical Aspects of Immunohistochemistry A ReviewDokument22 SeitenTechnical Aspects of Immunohistochemistry A Reviewacr3656Noch keine Bewertungen
- Anatomy and Physiology: Circulatory SystemDokument4 SeitenAnatomy and Physiology: Circulatory SystemJordz PlaciNoch keine Bewertungen
- (E BOOK) Martini Essentials of Anatomy PhysiologyDokument3 Seiten(E BOOK) Martini Essentials of Anatomy PhysiologyRaisa sya12Noch keine Bewertungen
- Objective: AB NODokument10 SeitenObjective: AB NOsamiNoch keine Bewertungen
- Personal Information Chief Complain History of Presenting IllnessDokument5 SeitenPersonal Information Chief Complain History of Presenting IllnessMuradNoch keine Bewertungen
- SNAP EquinosDokument24 SeitenSNAP Equinosapi-27393094Noch keine Bewertungen
- The Whole Is Greater Than The Sum of Its PartsDokument12 SeitenThe Whole Is Greater Than The Sum of Its PartsMabelPinesNoch keine Bewertungen
- Brain TumoursDokument35 SeitenBrain TumoursHealth Education Library for PeopleNoch keine Bewertungen
- Methyldopa DYNAMED 2014Dokument19 SeitenMethyldopa DYNAMED 2014Lalo AngelesNoch keine Bewertungen
- 4 - Ditta - BIO173 - Unity of Life - 20204Dokument13 Seiten4 - Ditta - BIO173 - Unity of Life - 20204John ChoiNoch keine Bewertungen
- Loco For Cocoa 1 Running Head: LOCO FOR COCOADokument13 SeitenLoco For Cocoa 1 Running Head: LOCO FOR COCOAapi-310439970Noch keine Bewertungen
- Midas VeniceDokument46 SeitenMidas Venicemirescualin100% (1)
- 86 Normal Low Tension Glaucoma PDFDokument4 Seiten86 Normal Low Tension Glaucoma PDFSherZalattha KuchikiElfNoch keine Bewertungen
- Training Effects of Long Versus Short Bouts of Exercise in Healthy Subjects.Dokument4 SeitenTraining Effects of Long Versus Short Bouts of Exercise in Healthy Subjects.HosseinNoch keine Bewertungen
- 67Dokument57 Seiten67Neeruam Ellehcim Aled ZurcNoch keine Bewertungen
- Enzyme Catalysis. FOXIT - Not Different, Just Better NAT91 Sin ReferenciasDokument6 SeitenEnzyme Catalysis. FOXIT - Not Different, Just Better NAT91 Sin Referenciaskiper.valNoch keine Bewertungen
- An Estimation of The Number of Cells in The Human BodyDokument11 SeitenAn Estimation of The Number of Cells in The Human BodysedilotNoch keine Bewertungen
- Final Poly 1Dokument44 SeitenFinal Poly 1kimberlyn odoñoNoch keine Bewertungen
- Chemistry 3.part 1pptxDokument62 SeitenChemistry 3.part 1pptxMaria Cyril DalusongNoch keine Bewertungen
- Histology Transes MidtermDokument13 SeitenHistology Transes MidtermblessganzoncandariNoch keine Bewertungen
- ALLOSTERISMDokument30 SeitenALLOSTERISMFlora D Souza100% (1)
- 1289 - Andrology Labratory ManualDokument245 Seiten1289 - Andrology Labratory ManualJuragang PotilNoch keine Bewertungen