Das könnte Ihnen auch gefallen
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5795)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Chrysler Dodge Ram Jeep Drive Cycle InformationDokument2 SeitenChrysler Dodge Ram Jeep Drive Cycle InformationslpkthNoch keine Bewertungen
- HLA HART Concept of LawDokument19 SeitenHLA HART Concept of LawHarneet KaurNoch keine Bewertungen
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Packing List For GermanyDokument2 SeitenPacking List For GermanyarjungangadharNoch keine Bewertungen
- Practical Research 2Dokument16 SeitenPractical Research 2Benedict Coloma BandongNoch keine Bewertungen
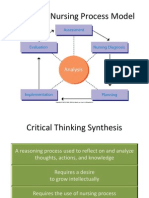
- NUR 104 Nursing Process MY NOTESDokument77 SeitenNUR 104 Nursing Process MY NOTESmeanne073100% (1)
- Revised LabDokument18 SeitenRevised LabAbu AyemanNoch keine Bewertungen
- English Lesson Plan Form 4 (Literature: "The Living Photograph")Dokument2 SeitenEnglish Lesson Plan Form 4 (Literature: "The Living Photograph")Maisarah Mohamad100% (3)
- Recruitment Process Outsourcing PDFDokument4 SeitenRecruitment Process Outsourcing PDFDevesh NamdeoNoch keine Bewertungen
- Self-Actualization in Robert Luketic'S: Legally Blonde: A HumanisticDokument10 SeitenSelf-Actualization in Robert Luketic'S: Legally Blonde: A HumanisticAyeshia FréyNoch keine Bewertungen
- NIH Public Access: Author ManuscriptDokument16 SeitenNIH Public Access: Author Manuscriptashgee1Noch keine Bewertungen
- Congenital Adrenal HyperplasiaDokument13 SeitenCongenital Adrenal Hyperplasiaashgee1Noch keine Bewertungen
- THC Detection WindowDokument16 SeitenTHC Detection Windowashgee1Noch keine Bewertungen
- The Adrenal GlandsDokument34 SeitenThe Adrenal Glandsashgee1Noch keine Bewertungen
- A Simple Membrane Osmometer System & Experiments That Quantitatively Measure Osmotic PressureDokument9 SeitenA Simple Membrane Osmometer System & Experiments That Quantitatively Measure Osmotic Pressureashgee1Noch keine Bewertungen
- Diffusion Across A Sheep Red Blood Cell MembraneDokument26 SeitenDiffusion Across A Sheep Red Blood Cell MembraneGelo JosonNoch keine Bewertungen
- Carol Hanisch - The Personal Is PoliticalDokument5 SeitenCarol Hanisch - The Personal Is Politicalconradofalbo100% (1)
- The Hukbalahap InsurrectionDokument76 SeitenThe Hukbalahap Insurrectionashgee1Noch keine Bewertungen
- The Hukbalahap MovementDokument9 SeitenThe Hukbalahap Movementashgee1Noch keine Bewertungen
- Class Activity 1Dokument2 SeitenClass Activity 1ashgee1Noch keine Bewertungen
- CIT 811 TMA 4 Quiz QuestionDokument3 SeitenCIT 811 TMA 4 Quiz QuestionjohnNoch keine Bewertungen
- Liquid Process Piping - Part 1 General Piping Design PDFDokument33 SeitenLiquid Process Piping - Part 1 General Piping Design PDFnitin guptaNoch keine Bewertungen
- Livro Spectrochemical AnalysisDokument9 SeitenLivro Spectrochemical AnalysisRicardo PossebonNoch keine Bewertungen
- 56257375851Dokument3 Seiten56257375851Anneliese FernandesNoch keine Bewertungen
- House Staff OrderDokument2 SeitenHouse Staff OrderTarikNoch keine Bewertungen
- Differentiating Language Difference and Language Disorder - Information For Teachers Working With English Language Learners in The Schools PDFDokument23 SeitenDifferentiating Language Difference and Language Disorder - Information For Teachers Working With English Language Learners in The Schools PDFIqra HassanNoch keine Bewertungen
- Alfa Laval Aalborg Os Tci Marine BoilerDokument2 SeitenAlfa Laval Aalborg Os Tci Marine Boilera.lobanov2020Noch keine Bewertungen
- Ruby Onyinyechi Amanze - Werte Magazine - 2019Dokument2 SeitenRuby Onyinyechi Amanze - Werte Magazine - 2019José LaertonNoch keine Bewertungen
- TuberkulosisDokument285 SeitenTuberkulosisTeuku M. FebriansyahNoch keine Bewertungen
- Zoology LAB Scheme of Work 2023 Hsslive HSSDokument7 SeitenZoology LAB Scheme of Work 2023 Hsslive HSSspookyvibee666Noch keine Bewertungen
- Datasheet TBJ SBW13009-KDokument5 SeitenDatasheet TBJ SBW13009-KMarquinhosCostaNoch keine Bewertungen
- SBU PlanningDokument13 SeitenSBU PlanningMohammad Raihanul HasanNoch keine Bewertungen
- Electrostatics Practice ProblemsDokument4 SeitenElectrostatics Practice ProblemsMohammed Aftab AhmedNoch keine Bewertungen
- Case Study 17 TomDokument7 SeitenCase Study 17 Tomapi-519148723Noch keine Bewertungen
- Cisco Nexus 7000 Introduction To NX-OS Lab GuideDokument38 SeitenCisco Nexus 7000 Introduction To NX-OS Lab Guiderazzzzzzzzzzz100% (1)
- Cruiziat Et Al. 2002Dokument30 SeitenCruiziat Et Al. 2002Juan David TurriagoNoch keine Bewertungen
- LAB Find Out The Bottle ContentsDokument4 SeitenLAB Find Out The Bottle ContentschanNoch keine Bewertungen
- FBDokument27 SeitenFBBenjaminNoch keine Bewertungen
- Factsheet AMRT 2023 01Dokument4 SeitenFactsheet AMRT 2023 01arsyil1453Noch keine Bewertungen
- TML IML DefinitionDokument2 SeitenTML IML DefinitionFicticious UserNoch keine Bewertungen
- Procedure For Non Hazardous Waste ManagementDokument6 SeitenProcedure For Non Hazardous Waste ManagementPaddyNoch keine Bewertungen