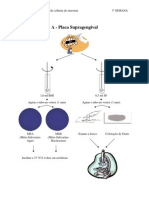
Das könnte Ihnen auch gefallen
- Liga MetálicaDokument9 SeitenLiga MetálicaAlexandra BatistaNoch keine Bewertungen
- Queda PDFDokument10 SeitenQueda PDFMarco AlvesNoch keine Bewertungen
- Ficha Formativa - Regulação GénicaDokument4 SeitenFicha Formativa - Regulação GénicaRubina FerreiraNoch keine Bewertungen
- Proposta de Exame Biologia e GeologiaDokument11 SeitenProposta de Exame Biologia e GeologiaCintia Brigita SilvaNoch keine Bewertungen
- Formação e crescimento de cristaisDokument21 SeitenFormação e crescimento de cristaisEduarda Cardozo100% (1)
- Solucoes Do Manual Biogeo 11 BioDokument34 SeitenSolucoes Do Manual Biogeo 11 BioCarina SilvaNoch keine Bewertungen
- Análise de Circuitos ElétricosDokument88 SeitenAnálise de Circuitos ElétricosBernardo CascãoNoch keine Bewertungen
- Aquela Triste e Leda MadrugadaDokument2 SeitenAquela Triste e Leda MadrugadamateusNoch keine Bewertungen
- Exercicios de BiologiaDokument9 SeitenExercicios de BiologiacarolNoch keine Bewertungen
- Questões AlsDokument8 SeitenQuestões AlsInês DominguezNoch keine Bewertungen
- 110Q - (Teste 2) - Nov2019Dokument5 Seiten110Q - (Teste 2) - Nov2019antonioNoch keine Bewertungen
- Destilação simples e fraccionadaDokument22 SeitenDestilação simples e fraccionadaCristiana SantosNoch keine Bewertungen
- Sistema reprodutor humano e desenvolvimento embrionárioDokument6 SeitenSistema reprodutor humano e desenvolvimento embrionáriocarolinaNoch keine Bewertungen
- Fatores EnzimaDokument5 SeitenFatores EnzimaEmily BarretoNoch keine Bewertungen
- Desenvolvimento e Custos Ecológicos Catarina Ferreira (1) (2385)Dokument9 SeitenDesenvolvimento e Custos Ecológicos Catarina Ferreira (1) (2385)Camila Soares100% (1)
- Poster - Um Pesadelo Chamado InfertilidadeDokument1 SeitePoster - Um Pesadelo Chamado InfertilidadeBernadetteNoch keine Bewertungen
- Soluções Livro 11 AnoDokument81 SeitenSoluções Livro 11 AnoOBerçodaCortiçaNoch keine Bewertungen
- T 12A Bio2 15Dokument5 SeitenT 12A Bio2 15Maria MaiaNoch keine Bewertungen
- Sebenta FinalDokument740 SeitenSebenta FinalRodrigo FernandesNoch keine Bewertungen
- Poster Da Infertilidade Final Grupo 1 Turno 2Dokument1 SeitePoster Da Infertilidade Final Grupo 1 Turno 2ruisoares65100% (1)
- Autofagia descobertaDokument4 SeitenAutofagia descobertaSandraNoch keine Bewertungen
- Relatório 3Dokument7 SeitenRelatório 3António LucasNoch keine Bewertungen
- ArealDokument12 SeitenArealLuísa PintoNoch keine Bewertungen
- Probabilidade III - Matematica 12ºano - PTDokument19 SeitenProbabilidade III - Matematica 12ºano - PTPedro SantosNoch keine Bewertungen
- Fisiologia RenalDokument14 SeitenFisiologia RenalCristiana PachecoNoch keine Bewertungen
- Atrito estático e cinético: registos e resultados experimentaisDokument15 SeitenAtrito estático e cinético: registos e resultados experimentaisSofia RodriguesNoch keine Bewertungen
- Q12 - TesteDokument8 SeitenQ12 - TesteCarlos PocinhoNoch keine Bewertungen
- Fermentação LevedurasDokument16 SeitenFermentação LevedurascarolinaNoch keine Bewertungen
- Bio12 Ta1Dokument6 SeitenBio12 Ta1Mariana GuimarãesNoch keine Bewertungen
- Novo11Q Critérios Específicos Classificação Teste3 Mai18Dokument3 SeitenNovo11Q Critérios Específicos Classificação Teste3 Mai18pituchaNoch keine Bewertungen
- Edicoes ASA - 11 Ano 2019-20 - 4 Teste PDFDokument9 SeitenEdicoes ASA - 11 Ano 2019-20 - 4 Teste PDFrefúgio do saberNoch keine Bewertungen
- Solubilidade de sais e iões na água do marDokument7 SeitenSolubilidade de sais e iões na água do marOrlando Meireles0% (1)
- Ondas mecânicas e acústicas em diferentes meiosDokument2 SeitenOndas mecânicas e acústicas em diferentes meiostita32Noch keine Bewertungen
- Apresentação - Deformação Das RochasDokument31 SeitenApresentação - Deformação Das RochasBia BorgesNoch keine Bewertungen
- Complexos e CorDokument15 SeitenComplexos e CorFilipe LopesNoch keine Bewertungen
- Teste Matemática Porto Turistas 2022Dokument3 SeitenTeste Matemática Porto Turistas 2022Bia sousa0% (1)
- Relatório Biologia - Transporte Nas Plantas PDFDokument30 SeitenRelatório Biologia - Transporte Nas Plantas PDFO Sérgio ConceiçãoNoch keine Bewertungen
- Encontros Português 11o AnoDokument9 SeitenEncontros Português 11o AnoAna Isabel Lopes Marques 1Noch keine Bewertungen
- Atividade Experimental Coeficiente de Expansao LinearDokument10 SeitenAtividade Experimental Coeficiente de Expansao LinearCarinaGasparNoch keine Bewertungen
- 11FQA Ficha Trab Ini F1.3 - N.º 4 CorrDokument6 Seiten11FQA Ficha Trab Ini F1.3 - N.º 4 CorrMaria Clara De Sá Bôtto100% (1)
- Sistema Excretor - 9ºanoDokument20 SeitenSistema Excretor - 9ºanoAna Sofia SantosNoch keine Bewertungen
- Energia térmica e radiação IIDokument7 SeitenEnergia térmica e radiação IIMaria DuarteNoch keine Bewertungen
- Novo 11Q NL (Teste2 CC) Abr.2019Dokument3 SeitenNovo 11Q NL (Teste2 CC) Abr.2019Rui SantosNoch keine Bewertungen
- Relatório 1Dokument14 SeitenRelatório 1Alexandra BalçaNoch keine Bewertungen
- Biologia 12.º AnoDokument65 SeitenBiologia 12.º AnoPaula Amado100% (1)
- Re82137 Ny12 Resolucoes Manual Vol1e2Dokument128 SeitenRe82137 Ny12 Resolucoes Manual Vol1e2Carina Cardoso CardosoNoch keine Bewertungen
- Sentidos 11Dokument1 SeiteSentidos 11Miguel Dias100% (1)
- Novo11Q - (Teste 3 - Mai18)Dokument5 SeitenNovo11Q - (Teste 3 - Mai18)Isabel Vilhena FerreiraNoch keine Bewertungen
- V Gowin Sedimentacao 10ºano de Escolaridafwcome Trhcejveby YvyhDokument1 SeiteV Gowin Sedimentacao 10ºano de Escolaridafwcome Trhcejveby YvyhFly SweetNoch keine Bewertungen
- Teste 1 Fisica 12 Ano Novembro 2021 Turma ADokument3 SeitenTeste 1 Fisica 12 Ano Novembro 2021 Turma ADiretor do AgrupamentoNoch keine Bewertungen
- Aef11 Teste Ava 1Dokument5 SeitenAef11 Teste Ava 1TeresaVaraoNoch keine Bewertungen
- Ensaio FilosoficoDokument2 SeitenEnsaio FilosoficoRafaela Carvalho Alunos AECANoch keine Bewertungen
- Fq10a BQ 00010Dokument6 SeitenFq10a BQ 00010Maria DuarteNoch keine Bewertungen
- Apontamentos de Biologia sobre o Sistema Reprodutor MasculinoDokument93 SeitenApontamentos de Biologia sobre o Sistema Reprodutor MasculinoAna Rita EstevesNoch keine Bewertungen
- Aplicações Informáticas - Introdução ProgramaçãoDokument6 SeitenAplicações Informáticas - Introdução ProgramaçãoSofia RochaNoch keine Bewertungen
- Atividade Laboratorial Nº 3 - Produção de Iogurte Natural PDFDokument1 SeiteAtividade Laboratorial Nº 3 - Produção de Iogurte Natural PDFJosé Luiz Rodrigues SilvaNoch keine Bewertungen
- Resumo Tecido Muscular - JunqueiraDokument4 SeitenResumo Tecido Muscular - JunqueiraLucas Thiago BorgesNoch keine Bewertungen
- Tecido MuscularDokument4 SeitenTecido MuscularDeadmanNoch keine Bewertungen
- Fisiologia Av 2 k4zDokument89 SeitenFisiologia Av 2 k4zCaroline MoreiraNoch keine Bewertungen
- Medresumos 2016 - OmfDokument16 SeitenMedresumos 2016 - OmfMirianNoch keine Bewertungen
- PROGORALDokument36 SeitenPROGORALapi-26429188Noch keine Bewertungen
- OralDokument54 SeitenOralapi-26429188100% (2)
- Microbiologia Oral Microbiologia Oral ApontamentosDokument6 SeitenMicrobiologia Oral Microbiologia Oral Apontamentosapi-26429188100% (2)
- 2 Frekmicrobiologia2005Dokument1 Seite2 Frekmicrobiologia2005api-26429188Noch keine Bewertungen
- 2 Frekmicrobiologia2005Dokument1 Seite2 Frekmicrobiologia2005api-26429188Noch keine Bewertungen
- Examemicrobiologiajulho 2005Dokument1 SeiteExamemicrobiologiajulho 2005api-26429188Noch keine Bewertungen
- HepatiteDokument3 SeitenHepatiteapi-26429188Noch keine Bewertungen
- Microbiologia Oral Esquema Protocolo 10 - SEMANA API20ADokument7 SeitenMicrobiologia Oral Esquema Protocolo 10 - SEMANA API20Aapi-26429188Noch keine Bewertungen
- Microbiologia Oral 3 AULA TPDokument13 SeitenMicrobiologia Oral 3 AULA TPapi-26429188100% (1)
- Doenças PeriodontaisDokument6 SeitenDoenças Periodontaisapi-26429188100% (3)
- FW Micro Oral Micro-2 - FreqDokument1 SeiteFW Micro Oral Micro-2 - Freqapi-26429188Noch keine Bewertungen
- HepatiteDokument3 SeitenHepatiteapi-26429188Noch keine Bewertungen
- Exame de Micro Geral - Fevereiro de 2005 MDDokument8 SeitenExame de Micro Geral - Fevereiro de 2005 MDapi-26429188Noch keine Bewertungen
- Ex Ames 1Dokument13 SeitenEx Ames 1api-26429188Noch keine Bewertungen
- Esquema Protocolo 4 SEMANADokument3 SeitenEsquema Protocolo 4 SEMANAapi-26429188Noch keine Bewertungen
- Esquema Protocolo 5 SEMANADokument7 SeitenEsquema Protocolo 5 SEMANAapi-26429188Noch keine Bewertungen
- Microbiologia Oral Esquema Protocolo 10 - SEMANA API20ADokument7 SeitenMicrobiologia Oral Esquema Protocolo 10 - SEMANA API20Aapi-26429188Noch keine Bewertungen
- Exame de Micro - Fev2004Dokument6 SeitenExame de Micro - Fev2004api-26429188Noch keine Bewertungen
- Esquema - Protocolo 3 SemanaDokument3 SeitenEsquema - Protocolo 3 Semanaapi-26429188Noch keine Bewertungen
- Escolha Multipla MICRO ORAL 1º Teste PraticoDokument2 SeitenEscolha Multipla MICRO ORAL 1º Teste Praticoapi-26429188Noch keine Bewertungen
- Doenças PeriodontaisDokument6 SeitenDoenças Periodontaisapi-26429188100% (3)
- Escolha Multipla MICRO ORAL 1º Teste PraticoDokument2 SeitenEscolha Multipla MICRO ORAL 1º Teste Praticoapi-26429188Noch keine Bewertungen
- Microbiologia Oral Microbiologia Oral ApontamentosDokument6 SeitenMicrobiologia Oral Microbiologia Oral Apontamentosapi-26429188100% (2)
- Escolha Multipla MICRO ORAL 1º Teste PraticoDokument2 SeitenEscolha Multipla MICRO ORAL 1º Teste Praticoapi-26429188Noch keine Bewertungen
- CarieDokument5 SeitenCarieapi-26429188Noch keine Bewertungen
- 2 Freq MicroDokument5 Seiten2 Freq Microapi-26429188Noch keine Bewertungen
- Doenças PeriodontaisDokument6 SeitenDoenças Periodontaisapi-26429188100% (3)
- BactériasDokument2 SeitenBactériasapi-26429188Noch keine Bewertungen
- 3 Aula TPDokument23 Seiten3 Aula TPapi-26429188Noch keine Bewertungen
- 2 Freq MicroDokument5 Seiten2 Freq Microapi-26429188Noch keine Bewertungen
- Exercicios para Desvios de JoelhoDokument5 SeitenExercicios para Desvios de JoelhoAndressa Souza100% (1)
- Skill TrainingDokument13 SeitenSkill TrainingDIOGO FRANCO PUREZANoch keine Bewertungen
- Assessoria Esportiva Twin: Treino ADokument4 SeitenAssessoria Esportiva Twin: Treino AJoão PauloNoch keine Bewertungen
- Revisão Músculos e ArticulaçõesDokument8 SeitenRevisão Músculos e ArticulaçõesLarissa SenaNoch keine Bewertungen
- Força muscular: avaliação e testesDokument52 SeitenForça muscular: avaliação e testesCaroline Menezes63% (8)
- Biomecânica Aula - Ligamentos e TendãoDokument28 SeitenBiomecânica Aula - Ligamentos e TendãoAnonymous LDLu9QrZd100% (2)
- Gestão equinoculturaDokument32 SeitenGestão equinoculturaJoao Francisco Pires LuizNoch keine Bewertungen
- Biofísica Músculo ContraçãoDokument30 SeitenBiofísica Músculo ContraçãoJô Joserlane Xavier100% (3)
- Contração MuscularDokument36 SeitenContração MuscularDiego FélixNoch keine Bewertungen
- Aula IV - Músculos Da Região Do QuadrilDokument18 SeitenAula IV - Músculos Da Região Do Quadrilapi-3729014100% (7)
- Ficha de Treino Klailson Robert Dos Santos CorreaDokument1 SeiteFicha de Treino Klailson Robert Dos Santos CorreaK Robert Correa100% (2)
- Aula de Miologia + RoteiroDokument36 SeitenAula de Miologia + RoteiroEdwin Te vendeNoch keine Bewertungen
- Joelho: Exame Físico CompletoDokument5 SeitenJoelho: Exame Físico CompletoLucas TelesNoch keine Bewertungen
- Importância dos ossos, articulações e músculosDokument30 SeitenImportância dos ossos, articulações e músculosmldexterNoch keine Bewertungen
- 5 MiologiaDokument72 Seiten5 MiologiasgtwelberNoch keine Bewertungen
- Resumo Tecido Muscular - JunqueiraDokument4 SeitenResumo Tecido Muscular - JunqueiraLucas Thiago BorgesNoch keine Bewertungen
- Controle Neuromuscular Do MovimentoDokument45 SeitenControle Neuromuscular Do MovimentoNavegantes Silva de OliveiraNoch keine Bewertungen
- Aula Cotovelo - Punho - MaoDokument87 SeitenAula Cotovelo - Punho - MaoSuely Suzuki VeraNoch keine Bewertungen
- Seção+2 4+-+sistema+muscular+origem,+organização+geral+e+histologiaDokument23 SeitenSeção+2 4+-+sistema+muscular+origem,+organização+geral+e+histologiamegamenteNoch keine Bewertungen
- Treino musculação intermediáriosDokument18 SeitenTreino musculação intermediáriosMarius ViníciusNoch keine Bewertungen
- Anatomia e Fisiologia do Sistema MuscularDokument27 SeitenAnatomia e Fisiologia do Sistema MuscularAdão 21Noch keine Bewertungen
- Atividade de Aprendizagem. MuscularDokument17 SeitenAtividade de Aprendizagem. Muscularmarcosbigandrade89% (9)
- Resumo do Aparelho LocomotorDokument6 SeitenResumo do Aparelho LocomotorCaroline LimaNoch keine Bewertungen
- Alongamento Excêntrico X Alongamento PassivoDokument9 SeitenAlongamento Excêntrico X Alongamento PassivoGabriel LopesNoch keine Bewertungen
- Sistema muscular: propriedades, tipos e ação dos músculosDokument2 SeitenSistema muscular: propriedades, tipos e ação dos músculosmaria eduardaNoch keine Bewertungen
- Biomecânica Da Cintura Escapular e OmbroDokument7 SeitenBiomecânica Da Cintura Escapular e OmbroMagnunLanfranco100% (2)
- Tratamento não cirúrgico da dor femoropatelarDokument2 SeitenTratamento não cirúrgico da dor femoropatelarTainá OrestesNoch keine Bewertungen
- Biomecanica Do Tornozelo e PeDokument39 SeitenBiomecanica Do Tornozelo e PeArquivos de Fisioterapia100% (4)
- Tecido MuscularDokument62 SeitenTecido MuscularThiago Pimenta100% (1)
- Treino ABC 2X AdaptaçãoDokument6 SeitenTreino ABC 2X AdaptaçãobambambigelowNoch keine Bewertungen