Das könnte Ihnen auch gefallen
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Anatomy and Physiology Cardiovascular System The HeartDokument6 SeitenAnatomy and Physiology Cardiovascular System The Heartmark OrpillaNoch keine Bewertungen
- Heal Yourself Naturally PaidalajinDokument219 SeitenHeal Yourself Naturally Paidalajinluis alejandro osorio100% (7)
- 7th Grade Natural Science Reinforcement WorksheetDokument3 Seiten7th Grade Natural Science Reinforcement WorksheetnataliaNoch keine Bewertungen
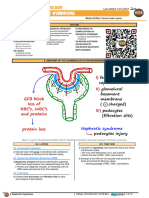
- Renal Pathology - 012) Nephrotic Syndrome (Notes)Dokument10 SeitenRenal Pathology - 012) Nephrotic Syndrome (Notes)hasanatiya41Noch keine Bewertungen
- The Kidney in Normal Aging: A Comparison With Chronic Kidney DiseaseDokument3 SeitenThe Kidney in Normal Aging: A Comparison With Chronic Kidney DiseasePorfirio Astudillo CastroNoch keine Bewertungen
- G Hargrove Hybrid DialysisDokument20 SeitenG Hargrove Hybrid DialysisDewiyanti TodingNoch keine Bewertungen
- Assessment of The Renal and Urinary Tract Syatems NewDokument27 SeitenAssessment of The Renal and Urinary Tract Syatems NewMay LindaNoch keine Bewertungen
- Grade 11 Life Sciences Revision Material Term 3 and 4 - 2023Dokument41 SeitenGrade 11 Life Sciences Revision Material Term 3 and 4 - 2023Junior NgwenyaNoch keine Bewertungen
- Chronic Kidney Disease Case PresentationDokument14 SeitenChronic Kidney Disease Case PresentationEmi EspinoNoch keine Bewertungen
- (Jurnal) Palliative and End-Of-life Care Issues in Chronic Kidney DiseaseDokument10 Seiten(Jurnal) Palliative and End-Of-life Care Issues in Chronic Kidney DiseaseRizqi DifaNoch keine Bewertungen
- Resuscitation Fluid Composition in Acute Kidney Injury in Critical IllnessDokument2 SeitenResuscitation Fluid Composition in Acute Kidney Injury in Critical IllnessDe Frede EditoreNoch keine Bewertungen
- Características Ecógrafo REPRO 4D SIUI CTS 800Dokument6 SeitenCaracterísticas Ecógrafo REPRO 4D SIUI CTS 800Cuarta Dimensión MedicaNoch keine Bewertungen
- Urinary SystemDokument80 SeitenUrinary SystemRoeisa Salinas100% (1)
- Surgery Student Logbook: Kasr Alainy Faculty of Medicine Cairo UniversityDokument40 SeitenSurgery Student Logbook: Kasr Alainy Faculty of Medicine Cairo UniversityGultach MatiyevaNoch keine Bewertungen
- Daftar Pustaka Radio JadiDokument2 SeitenDaftar Pustaka Radio JadiSartika AkibNoch keine Bewertungen
- Function of KidneyDokument66 SeitenFunction of KidneyNilesh100% (4)
- The Dates and Lemon Detox Protocol Sheet Official V-1.2Dokument20 SeitenThe Dates and Lemon Detox Protocol Sheet Official V-1.2Shaun Oneill100% (1)
- Hisam Nurokhman.a (201902010094)Dokument22 SeitenHisam Nurokhman.a (201902010094)Hisam hoboNoch keine Bewertungen
- Ingles StudyDokument23 SeitenIngles StudyMatiasUrraNoch keine Bewertungen
- Abortifacient and ContraceptivesDokument100 SeitenAbortifacient and ContraceptivesMario MorenoNoch keine Bewertungen
- Embryology of The KidneysDokument18 SeitenEmbryology of The KidneysPrincewill SmithNoch keine Bewertungen
- Caso ClinicoDokument4 SeitenCaso ClinicoNath ZarumaNoch keine Bewertungen
- Physiology - Guyton BLOODDokument6 SeitenPhysiology - Guyton BLOODIsra K. SoomroNoch keine Bewertungen
- KIDNEY DISEASE A HOLISTIC THERAPY by Walter LastDokument13 SeitenKIDNEY DISEASE A HOLISTIC THERAPY by Walter Lastmike davis100% (1)
- Biology 12: Provincial ExaminationDokument36 SeitenBiology 12: Provincial ExaminationVaniseNoch keine Bewertungen
- Aubf BrunzelDokument42 SeitenAubf Brunzelabigail lausNoch keine Bewertungen
- Renal Cell Carcinoma - Nursing Considerations With The Use of Targeted TherapyDokument50 SeitenRenal Cell Carcinoma - Nursing Considerations With The Use of Targeted TherapyMarivic DianoNoch keine Bewertungen
- Excretion in Humans 2 QPDokument18 SeitenExcretion in Humans 2 QPAhmad UmarNoch keine Bewertungen
- DLL Quarter 2 Week 1 (Melc - Idea - 5e's - Pivot - Adm-Pasay)Dokument64 SeitenDLL Quarter 2 Week 1 (Melc - Idea - 5e's - Pivot - Adm-Pasay)Misha Madeleine GacadNoch keine Bewertungen
- Forxiga Presentation Kaltim - Prof AgungDokument36 SeitenForxiga Presentation Kaltim - Prof AgungSanrio NeuroNoch keine Bewertungen