Das könnte Ihnen auch gefallen
- Gang Hu Et Al - Controlling Spiral Waves in A Model of Two-Dimensional Arrays of Chua's CircuitsDokument4 SeitenGang Hu Et Al - Controlling Spiral Waves in A Model of Two-Dimensional Arrays of Chua's CircuitsGretymjNoch keine Bewertungen
- Georg Gottwald, Alain Pumir and Valentin Krinsky - Spiral Wave Drift Induced by Stimulating Wave TrainsDokument8 SeitenGeorg Gottwald, Alain Pumir and Valentin Krinsky - Spiral Wave Drift Induced by Stimulating Wave TrainsGretymjNoch keine Bewertungen
- James Kent - Toward A Unified Theory of Psychedelic Action Or: "Hallucinogens and Recurrent Excitation in Cortical Circuitry"Dokument1 SeiteJames Kent - Toward A Unified Theory of Psychedelic Action Or: "Hallucinogens and Recurrent Excitation in Cortical Circuitry"GretymjNoch keine Bewertungen
- Yuan Xiao-Ping Et Al - Spiral Wave Generation in A Vortex Electric FieldDokument4 SeitenYuan Xiao-Ping Et Al - Spiral Wave Generation in A Vortex Electric FieldGretymjNoch keine Bewertungen
- Ma Jun Et Al - Breakup of Spiral Waves in Coupled Hindmarsh-Rose NeuronsDokument4 SeitenMa Jun Et Al - Breakup of Spiral Waves in Coupled Hindmarsh-Rose NeuronsGretymjNoch keine Bewertungen
- Jun Ma Et Al - The Instability of The Spiral Wave Induced by The Deformation of Elastic Excitable MediaDokument11 SeitenJun Ma Et Al - The Instability of The Spiral Wave Induced by The Deformation of Elastic Excitable MediaGretymjNoch keine Bewertungen
- KURENAI: Kyoto University Research Information RepositoryDokument5 SeitenKURENAI: Kyoto University Research Information RepositoryGretymjNoch keine Bewertungen
- Removal of A Pinned Spiral by Generating Target Waves With A Localized StimulusDokument5 SeitenRemoval of A Pinned Spiral by Generating Target Waves With A Localized StimulusGretymjNoch keine Bewertungen
- Michael Jachan Et Al - Inferring Direct Directed-Information Flow From Multivariate Nonlinear Time SeriesDokument5 SeitenMichael Jachan Et Al - Inferring Direct Directed-Information Flow From Multivariate Nonlinear Time SeriesGretymjNoch keine Bewertungen
- Complex-Periodic Spiral Waves in Confluent Cardiac Cell Cultures Induced by Localized InhomogeneitiesDokument7 SeitenComplex-Periodic Spiral Waves in Confluent Cardiac Cell Cultures Induced by Localized InhomogeneitiesGretymjNoch keine Bewertungen
- Marcel Hörning Et Al - Wave Emission On Interacting Heterogeneities in Cardiac TissueDokument8 SeitenMarcel Hörning Et Al - Wave Emission On Interacting Heterogeneities in Cardiac TissueGretymjNoch keine Bewertungen
- Zhoujian Cao Et Al - Turbulence Control With Local Pacing and Its Implication in Cardiac DefibrillationDokument9 SeitenZhoujian Cao Et Al - Turbulence Control With Local Pacing and Its Implication in Cardiac DefibrillationGretymjNoch keine Bewertungen
- Hong Zhang Et Al - Control of Spiral Breakup by An Alternating Advective FieldDokument5 SeitenHong Zhang Et Al - Control of Spiral Breakup by An Alternating Advective FieldGretymjNoch keine Bewertungen
- Sitabhra Sinha Et Al - Critical Role of Inhomogeneities in Pacing Termination of Cardiac ReentryDokument10 SeitenSitabhra Sinha Et Al - Critical Role of Inhomogeneities in Pacing Termination of Cardiac ReentryGretymjNoch keine Bewertungen
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Muscular SystemDokument43 SeitenMuscular SystemJonalyn ChewacheoNoch keine Bewertungen
- Cell - The Unit of Life - NCERT Based PYQsDokument9 SeitenCell - The Unit of Life - NCERT Based PYQsAkhil singhNoch keine Bewertungen
- Part of BodyDokument4 SeitenPart of Bodyanon_535068952Noch keine Bewertungen
- Microteaching Pituitary GLANDDokument6 SeitenMicroteaching Pituitary GLANDaparnaNoch keine Bewertungen
- Audience AnalysisDokument14 SeitenAudience AnalysisMarimar De Luna AuditorNoch keine Bewertungen
- Physiological Changes During Pregnancy 2-1Dokument32 SeitenPhysiological Changes During Pregnancy 2-1PATRICK ROSHAN ANoch keine Bewertungen
- BIBLIOGRAFIE Neural Conduction Synaptic TransmisionDokument16 SeitenBIBLIOGRAFIE Neural Conduction Synaptic TransmisionMirela GiurgeaNoch keine Bewertungen
- Cardiorespiratory SystemDokument15 SeitenCardiorespiratory SystemDafchen Villarin MahasolNoch keine Bewertungen
- Nervous System Mind MapDokument1 SeiteNervous System Mind MapAra Malagón González100% (1)
- DIGESTIVE Lecture Notes PDFDokument20 SeitenDIGESTIVE Lecture Notes PDFAaliya Minar100% (4)
- Skull, Brain and Orbit (Practical)Dokument45 SeitenSkull, Brain and Orbit (Practical)Chris Jardine LiNoch keine Bewertungen
- Lesson 5 Connective TissueDokument5 SeitenLesson 5 Connective Tissueshairene PiaNoch keine Bewertungen
- Audition: Hearing, The Ear, and Sound LocalizationDokument5 SeitenAudition: Hearing, The Ear, and Sound LocalizationTanishqa RawlaniNoch keine Bewertungen
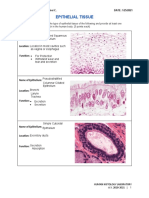
- Epithelial Tissue: Name of Epithelium: Stratified SquamousDokument3 SeitenEpithelial Tissue: Name of Epithelium: Stratified SquamousNoriel LacsinaNoch keine Bewertungen
- Erythema MultiformeDokument5 SeitenErythema MultiformeNicco MarantsonNoch keine Bewertungen
- A.D. Pearle - Surgical Technique and Anatomic Study of Latissimus Dorsi and Teres Major Transfers (2006)Dokument9 SeitenA.D. Pearle - Surgical Technique and Anatomic Study of Latissimus Dorsi and Teres Major Transfers (2006)João Pedro ZenattoNoch keine Bewertungen
- Trends in Neural EvolutionDokument35 SeitenTrends in Neural EvolutionTejpal Dahiya100% (1)
- Anatomy of Flowering Plants Mind MapDokument2 SeitenAnatomy of Flowering Plants Mind MapAstha Agrawal100% (8)
- The - Skeletal - System Lab ActDokument13 SeitenThe - Skeletal - System Lab Actwelfred indinoNoch keine Bewertungen
- Marieb Ch11aDokument23 SeitenMarieb Ch11aapi-229554503Noch keine Bewertungen
- Pediatrics CLINICAL QUESTIONDokument14 SeitenPediatrics CLINICAL QUESTIONAyesha KhatunNoch keine Bewertungen
- Extracellular VesicleDokument14 SeitenExtracellular VesicleLucas Monteiro LopesNoch keine Bewertungen
- Amniotic Fluid 2022Dokument40 SeitenAmniotic Fluid 2022Ogbuefi PascalNoch keine Bewertungen
- IGCSE Bio - Blood DonationDokument13 SeitenIGCSE Bio - Blood DonationTroy CampbellNoch keine Bewertungen
- Placenta PreviaDokument19 SeitenPlacenta PreviapriyankaNoch keine Bewertungen
- WORKSHEET 2.2 Cell Structure and FunctionDokument6 SeitenWORKSHEET 2.2 Cell Structure and FunctionasyuraNoch keine Bewertungen
- Periodontics PDFDokument28 SeitenPeriodontics PDFtriciaNoch keine Bewertungen
- Reproduction MCQssDokument6 SeitenReproduction MCQssPamela CastilloNoch keine Bewertungen
- Worksheet For Kinder - q1w6Dokument20 SeitenWorksheet For Kinder - q1w6lccyu9Noch keine Bewertungen