Das könnte Ihnen auch gefallen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (894)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (587)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (119)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2219)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Maximizing Growth Factors for Height IncreaseDokument29 SeitenMaximizing Growth Factors for Height IncreaseMegaV100% (2)
- Ron Kurtz - The Body Reveals PDFDokument161 SeitenRon Kurtz - The Body Reveals PDFNemanja92% (12)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Come Thirsty Chapter1Dokument4 SeitenCome Thirsty Chapter1Natalia Lupasco100% (1)
- Brainstem Bravo AnnotatedDokument13 SeitenBrainstem Bravo AnnotatedMia CadizNoch keine Bewertungen
- CONSUMER BEHAVIOUR UpdatedDokument66 SeitenCONSUMER BEHAVIOUR UpdatedUmer AzizNoch keine Bewertungen
- Abbreviation For ChartingDokument12 SeitenAbbreviation For ChartingJomar De BelenNoch keine Bewertungen
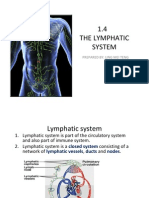
- 1.4 The Lymphatic System: Prepared By: Ling Mei TengDokument15 Seiten1.4 The Lymphatic System: Prepared By: Ling Mei TengJuliet LingNoch keine Bewertungen
- Risk For Decreased Cardiac OutputDokument4 SeitenRisk For Decreased Cardiac Outputapi-283482759Noch keine Bewertungen
- Biochem QbankDokument16 SeitenBiochem Qbank786waqar786Noch keine Bewertungen
- UntitledDokument89 SeitenUntitledVladimir VešovićNoch keine Bewertungen
- Amino Acid Sequences Reveal Evolutionary RelationshipsDokument6 SeitenAmino Acid Sequences Reveal Evolutionary RelationshipsAbby Gail DigalNoch keine Bewertungen
- SP FinalDokument18 SeitenSP Finalapi-344954734Noch keine Bewertungen
- Abdominal Compartment SyndromeDokument15 SeitenAbdominal Compartment SyndromePierinaNoch keine Bewertungen
- Pathology - Cardiovascular SystemDokument17 SeitenPathology - Cardiovascular SystemNdegwa Jesse100% (2)
- NCP of CavDokument3 SeitenNCP of CavHenry Roque TagalagNoch keine Bewertungen
- Prokaryotes and Eukaryotes: Strategies and Successes: Michael CarlileDokument3 SeitenProkaryotes and Eukaryotes: Strategies and Successes: Michael CarlileNurul ShazwaniNoch keine Bewertungen
- 50 Ways To HealDokument12 Seiten50 Ways To HealBrooke PenaNoch keine Bewertungen
- Tetralogy of Fallot Everything You Wanted To Know But Were Afraid To AskDokument8 SeitenTetralogy of Fallot Everything You Wanted To Know But Were Afraid To AsksofiaNoch keine Bewertungen
- Overview of Complications Occurring in The Post-Anesthesia Care UnitDokument14 SeitenOverview of Complications Occurring in The Post-Anesthesia Care UnitShahabuddin ShaikhNoch keine Bewertungen
- Standardized Tools for Assessing Traumatic Brain InjuryDokument5 SeitenStandardized Tools for Assessing Traumatic Brain InjuryGenera Rose Radaza Villa100% (3)
- Pass MRCP Part 2 (Sample)Dokument26 SeitenPass MRCP Part 2 (Sample)Ahmad Shokry100% (1)
- Template POMRDokument8 SeitenTemplate POMRPPDS IPD ULMNoch keine Bewertungen
- 2018 Biology Exercises For SPM (Chapter6 & Chapter7)Dokument15 Seiten2018 Biology Exercises For SPM (Chapter6 & Chapter7)Kuen Jian LinNoch keine Bewertungen
- CorneaDokument41 SeitenCorneaNikhil KorripatiNoch keine Bewertungen
- Biology: Revised Syllabus For Higher Secondary First Year CourseDokument4 SeitenBiology: Revised Syllabus For Higher Secondary First Year CourseamitumikoibNoch keine Bewertungen
- NCM 103 RLE NOTESDokument8 SeitenNCM 103 RLE NOTESgallardo.bettinarose.iNoch keine Bewertungen
- Nilai NormalDokument5 SeitenNilai NormalAgya Ghilman FazaNoch keine Bewertungen
- Bacillus lipopeptides: versatile weapons against plant diseasesDokument11 SeitenBacillus lipopeptides: versatile weapons against plant diseasesjazminNoch keine Bewertungen
- Neurotransmission, Measuring Chemical Events In: Advanced ArticleDokument12 SeitenNeurotransmission, Measuring Chemical Events In: Advanced ArticleazzaassNoch keine Bewertungen
- Basic Anatomical Terms PPTDokument30 SeitenBasic Anatomical Terms PPTInsatiable Clee100% (1)