Das könnte Ihnen auch gefallen
- Lab Report Animal and Plant CellsDokument5 SeitenLab Report Animal and Plant CellsNicolas Gómez100% (1)
- Update in TBC and Non TBCDokument5 SeitenUpdate in TBC and Non TBCReynaVG19Noch keine Bewertungen
- Journal Ppat 1003468Dokument15 SeitenJournal Ppat 1003468Zoom TecnologicoNoch keine Bewertungen
- Plasma Histamine and Tumour Necrosis Factor-Alpha Levels in Crohn'S Disease and Ulcerative Colitis at Various Stages of DiseaseDokument8 SeitenPlasma Histamine and Tumour Necrosis Factor-Alpha Levels in Crohn'S Disease and Ulcerative Colitis at Various Stages of DiseaseNejc KovačNoch keine Bewertungen
- Prevalence of Helicobacter Pylori Infection and Its Association With Cardiovascular Risk Factors in Korean AdultsDokument7 SeitenPrevalence of Helicobacter Pylori Infection and Its Association With Cardiovascular Risk Factors in Korean AdultsAlbert PapilayaNoch keine Bewertungen
- Nihms 1688237Dokument20 SeitenNihms 1688237Mohammed KhalidNoch keine Bewertungen
- Systems Immunology of Diabetes-Tuberculosis Comorbidity Reveals Signatures of Disease ComplicationsDokument16 SeitenSystems Immunology of Diabetes-Tuberculosis Comorbidity Reveals Signatures of Disease Complicationsaini nazilaNoch keine Bewertungen
- DM1 Nature 2021Dokument20 SeitenDM1 Nature 2021Beatriz MDNoch keine Bewertungen
- P 219 HTLV 1 Infected Regulatory T Cell Expansion.255 PDFDokument2 SeitenP 219 HTLV 1 Infected Regulatory T Cell Expansion.255 PDFNilo BarretoNoch keine Bewertungen
- Inflammatorybowel Disease: Anne Walsh,, John Mabee,, Kashyap TrivediDokument18 SeitenInflammatorybowel Disease: Anne Walsh,, John Mabee,, Kashyap Trivediangeldavid26Noch keine Bewertungen
- Stüber F, 2004Dokument6 SeitenStüber F, 2004Hendry SusantoNoch keine Bewertungen
- Enfermedad de CrohnDokument16 SeitenEnfermedad de CrohnFabrizzio BardalesNoch keine Bewertungen
- Clinical Features and Major Histocompatibility Complex Genes As Potential Susceptibility Factors in Pediatric Immune ThrombocytopeniaDokument24 SeitenClinical Features and Major Histocompatibility Complex Genes As Potential Susceptibility Factors in Pediatric Immune ThrombocytopeniaMerlin Margreth MaelissaNoch keine Bewertungen
- TB and Host Metabolism 1Dokument8 SeitenTB and Host Metabolism 1Roger CNoch keine Bewertungen
- Infections As A Cause of Autoimmune Rheumatic Diseases.: ResumenDokument16 SeitenInfections As A Cause of Autoimmune Rheumatic Diseases.: ResumenSergio Revert BodíNoch keine Bewertungen
- Risk Factors For Extra-Pulmonary Tuberculosis Compared To Pulmonary TuberculosisDokument6 SeitenRisk Factors For Extra-Pulmonary Tuberculosis Compared To Pulmonary TuberculosisLily SmithNoch keine Bewertungen
- Relationship of Hla-Dw3 HLA-B8 To Sjogren'S Syndrome: (P (P F PDokument6 SeitenRelationship of Hla-Dw3 HLA-B8 To Sjogren'S Syndrome: (P (P F PMohmed AttiaNoch keine Bewertungen
- Tropmed 86 698 PDFDokument5 SeitenTropmed 86 698 PDFsandraldiraNoch keine Bewertungen
- ContentServer - Asp 11Dokument7 SeitenContentServer - Asp 11Fitria NurulfathNoch keine Bewertungen
- Differences in Clinical and Laboratory Findings Between Group D and Non-Group D Non-TyphoidalDokument10 SeitenDifferences in Clinical and Laboratory Findings Between Group D and Non-Group D Non-TyphoidalBana NanaNoch keine Bewertungen
- CISH and Susceptibility To Infectious Diseases: Original ArticleDokument10 SeitenCISH and Susceptibility To Infectious Diseases: Original ArticlenovianastasiaNoch keine Bewertungen
- Outcome and Clinical Characteristics in Pleural Empyema: A Retrospective StudyDokument7 SeitenOutcome and Clinical Characteristics in Pleural Empyema: A Retrospective StudylestarisurabayaNoch keine Bewertungen
- Pi Is 1083879112002820Dokument2 SeitenPi Is 1083879112002820ocampo silvinaNoch keine Bewertungen
- Pathophysiology of IBDDokument11 SeitenPathophysiology of IBDOktarina Heni SunandarNoch keine Bewertungen
- Healthmed 15 1 Damir SecicDokument6 SeitenHealthmed 15 1 Damir SecicEdoHNoch keine Bewertungen
- Ijerph 17 01742Dokument12 SeitenIjerph 17 01742journal proNoch keine Bewertungen
- Genomewide Association Study of Leprosy: Original ArticleDokument10 SeitenGenomewide Association Study of Leprosy: Original ArticleFickry AdiansyahNoch keine Bewertungen
- Ref 2Dokument9 SeitenRef 2indah setiyaniNoch keine Bewertungen
- Circulation ArticleDokument13 SeitenCirculation ArticleAjit JadhavNoch keine Bewertungen
- Wegener's Granulomatosis: Current and Upcoming Therapies: ReviewDokument12 SeitenWegener's Granulomatosis: Current and Upcoming Therapies: ReviewAfif FaiziNoch keine Bewertungen
- Fenotipurile Astmului 2008Dokument9 SeitenFenotipurile Astmului 2008childocNoch keine Bewertungen
- Nihms 822807Dokument15 SeitenNihms 822807Novita maraNoch keine Bewertungen
- Protective Effect of BCG Against Tuberculous Meningitis and Miliary Tuberculosis: A Meta-AnalysisDokument5 SeitenProtective Effect of BCG Against Tuberculous Meningitis and Miliary Tuberculosis: A Meta-AnalysisLely KarolinaNoch keine Bewertungen
- The Role of The Platelet-To-Lymphocyte Ratio and NDokument5 SeitenThe Role of The Platelet-To-Lymphocyte Ratio and NRudy Arindra WijayaNoch keine Bewertungen
- Sharma 2002Dokument4 SeitenSharma 2002wosen wondNoch keine Bewertungen
- Review: New Concepts in The Pathophysiology of Inflammatory Bowel DiseaseDokument11 SeitenReview: New Concepts in The Pathophysiology of Inflammatory Bowel DiseaseReynalth Andrew Sinaga100% (1)
- Fauzan Indra M. Lubis, Ricke Loesnihari, Taufik Sungkar: BackgroundDokument5 SeitenFauzan Indra M. Lubis, Ricke Loesnihari, Taufik Sungkar: BackgroundAl FatikhahNoch keine Bewertungen
- Aliment Pharmacol Ther - 2016 - Zhang - Letter Risk Factor and Mortality of Peptic Ulcer DiseaseDokument2 SeitenAliment Pharmacol Ther - 2016 - Zhang - Letter Risk Factor and Mortality of Peptic Ulcer DiseaseDrhussamNoch keine Bewertungen
- Immune Mechanisms in Type 1 Diabetes: Maja Wa Llberg and Anne CookeDokument9 SeitenImmune Mechanisms in Type 1 Diabetes: Maja Wa Llberg and Anne CookeNiko Hizkia SimatupangNoch keine Bewertungen
- Original Article: Assessment of A Lupus Nephritis Cohort Over A 30-Year PeriodDokument7 SeitenOriginal Article: Assessment of A Lupus Nephritis Cohort Over A 30-Year PerioddhineyNoch keine Bewertungen
- Association of The Interferon-C Gene (CA) Repeat Polymorphism With EndometriosisDokument6 SeitenAssociation of The Interferon-C Gene (CA) Repeat Polymorphism With EndometriosisGladstone AsadNoch keine Bewertungen
- Is There A Relationship Between Periodontal Disease and Causes of Death? A Cross Sectional StudyDokument6 SeitenIs There A Relationship Between Periodontal Disease and Causes of Death? A Cross Sectional StudyYaarit IustainNoch keine Bewertungen
- FOXP3 Variants Are Independently Associated With Transforming Growth Factor B1 Plasma Levels in Female Patients With Inflammatory Bowel DiseaseDokument9 SeitenFOXP3 Variants Are Independently Associated With Transforming Growth Factor B1 Plasma Levels in Female Patients With Inflammatory Bowel DiseasePaula KikuchiNoch keine Bewertungen
- Use of Flexible Bronchoscopy in The Diagnosis of Infectious Etiologies - A Literature ReviewDokument4 SeitenUse of Flexible Bronchoscopy in The Diagnosis of Infectious Etiologies - A Literature ReviewAna MercadoNoch keine Bewertungen
- ALSDokument14 SeitenALSZé CunhaNoch keine Bewertungen
- Neuro-Oncology and Palliative Care: A Challenging InterfaceDokument5 SeitenNeuro-Oncology and Palliative Care: A Challenging InterfaceJiaNoch keine Bewertungen
- Hep Chiv Renal DZDokument9 SeitenHep Chiv Renal DZSundar RamanathanNoch keine Bewertungen
- Genetics of Hypertension: The Lack of EvidenceDokument11 SeitenGenetics of Hypertension: The Lack of EvidenceInternational Medical PublisherNoch keine Bewertungen
- 539 FullDokument12 Seiten539 FullFadhel Jimmy AnugerahNoch keine Bewertungen
- Association Between Immune-Related Adverse Event TDokument8 SeitenAssociation Between Immune-Related Adverse Event TAndrew FincoNoch keine Bewertungen
- EditorialDokument2 SeitenEditorialsebastoto778Noch keine Bewertungen
- Ulcer and HelicobacterDokument10 SeitenUlcer and HelicobacterLiliana L SalazarNoch keine Bewertungen
- Genetic Investigationsof Meniere Sdisease: Jeffrey T. VrabecDokument12 SeitenGenetic Investigationsof Meniere Sdisease: Jeffrey T. Vrabecdisk_la_poduNoch keine Bewertungen
- 10 5588@ijtld 14 0963Dokument8 Seiten10 5588@ijtld 14 0963Oscar Rosero VasconesNoch keine Bewertungen
- Autoantibodies Against Complement C1q Correlate With The Thyroid Function in Patients With Autoimmune Thyroid DiseaseDokument6 SeitenAutoantibodies Against Complement C1q Correlate With The Thyroid Function in Patients With Autoimmune Thyroid DiseaseSaad MotawéaNoch keine Bewertungen
- HyperemesisDokument4 SeitenHyperemesisadindadian12Noch keine Bewertungen
- Jact 12 I 2 P 142Dokument3 SeitenJact 12 I 2 P 142Rosdiana Elizabeth Siburian0% (1)
- Possible Undertreatment of Women With Crohn Disease in PolandDokument7 SeitenPossible Undertreatment of Women With Crohn Disease in PolandFiraFurqaniNoch keine Bewertungen
- Which Patients Are Able To Adhere To Tuberculosis Treatment? A Study in A Rural Area in The Northwest Part of TurkeyDokument7 SeitenWhich Patients Are Able To Adhere To Tuberculosis Treatment? A Study in A Rural Area in The Northwest Part of TurkeyAnonymous QPXAgjBwNoch keine Bewertungen
- Disparities in Periodontitis Prevalence Among Chronic Kidney Disease PatientsDokument5 SeitenDisparities in Periodontitis Prevalence Among Chronic Kidney Disease PatientsKristelle HernandezNoch keine Bewertungen
- Auto-Inflammatory Syndromes: Pathophysiology, Diagnosis, and ManagementVon EverandAuto-Inflammatory Syndromes: Pathophysiology, Diagnosis, and ManagementPetros EfthimiouNoch keine Bewertungen
- 2018 CV PDF Dito AnurogoDokument15 Seiten2018 CV PDF Dito Anurogod17oNoch keine Bewertungen
- Your Electronic Ticket ReceiptDokument2 SeitenYour Electronic Ticket Receiptd17oNoch keine Bewertungen
- Fingerprint Prenatal DiagnosticDokument33 SeitenFingerprint Prenatal Diagnosticd17oNoch keine Bewertungen
- Childhood Traumas: An Outline and OverviewDokument13 SeitenChildhood Traumas: An Outline and Overviewd17o100% (1)
- Kacip Fatimah USA PatentDokument6 SeitenKacip Fatimah USA Patentd17oNoch keine Bewertungen
- The Prognosis of Breast Cancer in MalesDokument8 SeitenThe Prognosis of Breast Cancer in Malesd17oNoch keine Bewertungen
- Metabolites: An NMR Metabolomics Approach To Investigate Factors A and QualityDokument16 SeitenMetabolites: An NMR Metabolomics Approach To Investigate Factors A and QualitykarinaNoch keine Bewertungen
- 12480the Genesis of Gencidine PDFDokument6 Seiten12480the Genesis of Gencidine PDFPrasad KulkarniNoch keine Bewertungen
- Set A Suggested Answer Mosys SB025 - 230430 - 191516Dokument4 SeitenSet A Suggested Answer Mosys SB025 - 230430 - 191516nursyakiramailanNoch keine Bewertungen
- Quirino State University: Diffun CampusDokument2 SeitenQuirino State University: Diffun CampusMaden betoNoch keine Bewertungen
- Class Notes For Upper Biology MasterclassDokument2 SeitenClass Notes For Upper Biology MasterclassYehia Khaled Essam Fawaz ICSBR1Noch keine Bewertungen
- 2019 - Rol de La MITOCONDRIA Y PARKINSONDokument92 Seiten2019 - Rol de La MITOCONDRIA Y PARKINSONEduardo RodriguezNoch keine Bewertungen
- Bio - Cell Structure and Function QuizDokument2 SeitenBio - Cell Structure and Function QuizAditi AGRAWALNoch keine Bewertungen
- Proteomic Analysis of Coffee Grains Exposed To Different Drying ProcessDokument9 SeitenProteomic Analysis of Coffee Grains Exposed To Different Drying ProcessBeatriz MartinsNoch keine Bewertungen
- Immunofluorescence Handbook 1601971Dokument80 SeitenImmunofluorescence Handbook 1601971Ray KuoNoch keine Bewertungen
- Ligand ProteinDokument4 SeitenLigand ProteinOscar George LuNoch keine Bewertungen
- NFAT5 Amplifies Antipathogen Responses by Enhancing Chromatin Accessibility, H3K27 Demethylation, and Transcription Factor RecruitmentDokument17 SeitenNFAT5 Amplifies Antipathogen Responses by Enhancing Chromatin Accessibility, H3K27 Demethylation, and Transcription Factor RecruitmentFernandoNoch keine Bewertungen
- Amino AcidsDokument2 SeitenAmino AcidsAlliah MendozaNoch keine Bewertungen
- Bigdye Terminator V3.1 Cycle Sequencing Kit: ProtocolDokument72 SeitenBigdye Terminator V3.1 Cycle Sequencing Kit: Protocoliis faridahNoch keine Bewertungen
- Cia 2 347Dokument15 SeitenCia 2 347AnisNoch keine Bewertungen
- Bluher (2009) Adipose Tissue Dysfunction in ObesityDokument10 SeitenBluher (2009) Adipose Tissue Dysfunction in ObesityBruno TeixeiraNoch keine Bewertungen
- Chapter - 2 PDFDokument32 SeitenChapter - 2 PDFnikhilNoch keine Bewertungen
- Cse182 MT2022Dokument5 SeitenCse182 MT2022hylee102594Noch keine Bewertungen
- Anthrax Toxin Mechanism of ActionDokument8 SeitenAnthrax Toxin Mechanism of ActionWendy FXNoch keine Bewertungen
- Metabolism of XenobioticsDokument30 SeitenMetabolism of XenobioticsCzarina RiveraNoch keine Bewertungen
- Enzyme Temperature Lab ReportDokument5 SeitenEnzyme Temperature Lab Reportapi-401166686Noch keine Bewertungen
- CarbohydratesDokument31 SeitenCarbohydratesMarlene GonsalvezNoch keine Bewertungen
- Mechanism of Gastric SecretionDokument11 SeitenMechanism of Gastric Secretionmulya ningsihNoch keine Bewertungen
- 4 HMP ShuntDokument18 Seiten4 HMP ShuntAyoubMuhammadNoch keine Bewertungen
- DR Lal Pathlabs: LPL - Lpl-Rohini (National Reference Lab) Sector - 18, Block - E Rohini Delhi 110085Dokument4 SeitenDR Lal Pathlabs: LPL - Lpl-Rohini (National Reference Lab) Sector - 18, Block - E Rohini Delhi 110085Manojit SarkarNoch keine Bewertungen
- 2022 - Biochemistry - Esha D. Dalvie, Jordan C. Stacy, Keir C. Neuman - Recognition of DNA Supercoil Handedness During Catenation Catalyzed byDokument33 Seiten2022 - Biochemistry - Esha D. Dalvie, Jordan C. Stacy, Keir C. Neuman - Recognition of DNA Supercoil Handedness During Catenation Catalyzed byjimgogreatNoch keine Bewertungen
- Biological Science 6th Edition Freeman Test BankDokument22 SeitenBiological Science 6th Edition Freeman Test Bankscarletwilliamnfz100% (31)
- Lecture # 7 Pentose Phosphate PathwayDokument44 SeitenLecture # 7 Pentose Phosphate PathwayDesy AnggrainiNoch keine Bewertungen
- Nittophase HL Solid Support: High Loaded Polymeric Solid Supports For Oligonucleotide SynthesisDokument2 SeitenNittophase HL Solid Support: High Loaded Polymeric Solid Supports For Oligonucleotide Synthesiskasiman1234Noch keine Bewertungen
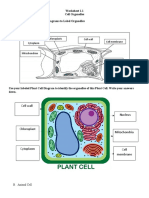
- Worksheet 1.1 Cell OrganelleDokument2 SeitenWorksheet 1.1 Cell OrganelleCyndel TindoyNoch keine Bewertungen