Das könnte Ihnen auch gefallen
- Mills The Comparative Anatomy of Eating1Dokument9 SeitenMills The Comparative Anatomy of Eating1acoohayNoch keine Bewertungen
- Department of Zoology University of Kashmir, Srinagar: by Prof. Fayaz AhmadDokument45 SeitenDepartment of Zoology University of Kashmir, Srinagar: by Prof. Fayaz Ahmadpre mdcatNoch keine Bewertungen
- Digestive Systems of Vertebrates and InvertebratesDokument7 SeitenDigestive Systems of Vertebrates and InvertebratesClexandrea Dela Luz CorpuzNoch keine Bewertungen
- Major Body Systems Flora: The Mouth and GI TractDokument2 SeitenMajor Body Systems Flora: The Mouth and GI TractAiza Mae Clacio LimNoch keine Bewertungen
- Growing Poultry Industry Meets Need for Alternative FeedsDokument47 SeitenGrowing Poultry Industry Meets Need for Alternative FeedsJakin Aia TapanganNoch keine Bewertungen
- Animal Digestive System Humms-11Dokument10 SeitenAnimal Digestive System Humms-11Culiao DelmarNoch keine Bewertungen
- Human Digestive System, The System Used in TheDokument10 SeitenHuman Digestive System, The System Used in Theعلي رسن رداد حمد ANoch keine Bewertungen
- TM 10 Only Small Animal and Monogastric Tract 25 July 2020 AGWDokument53 SeitenTM 10 Only Small Animal and Monogastric Tract 25 July 2020 AGWcellin rubianti azzharaNoch keine Bewertungen
- Comparative Vertebrate Anatomy of The Digestive System Source: Http://people - Eku.edu/ritchisong/342notes7.htmlDokument14 SeitenComparative Vertebrate Anatomy of The Digestive System Source: Http://people - Eku.edu/ritchisong/342notes7.htmlYeyen M. EvoraNoch keine Bewertungen
- The Comparative Anatomy of EatingDokument5 SeitenThe Comparative Anatomy of EatingBrian Wong100% (23)
- Digestive SystemDokument28 SeitenDigestive SystemChristian D. RamatNoch keine Bewertungen
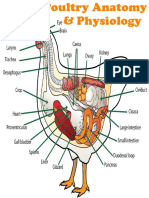
- Chicken Anatomy & PhysiologyDokument26 SeitenChicken Anatomy & PhysiologySanjeev ChaudharyNoch keine Bewertungen
- Comparative Anatomy: Table of ContentsDokument22 SeitenComparative Anatomy: Table of Contentsnutrientdense9183Noch keine Bewertungen
- Comparative Vertebrate Anatomy Lecture Notes - Digestive SystemDokument14 SeitenComparative Vertebrate Anatomy Lecture Notes - Digestive SystemMielah Ruth100% (1)
- Book 11 AnimalNutrition2021 Adapted 2Dokument11 SeitenBook 11 AnimalNutrition2021 Adapted 2assf7235Noch keine Bewertungen
- Lea 417 510 2011Dokument121 SeitenLea 417 510 2011Oliver TalipNoch keine Bewertungen
- Heterotrophic nutrition: how organisms obtain foodDokument10 SeitenHeterotrophic nutrition: how organisms obtain foodNakshatra PaliwalNoch keine Bewertungen
- MSF414 Term Paper - Evergreen OdehDokument6 SeitenMSF414 Term Paper - Evergreen OdehEvergreen OdehNoch keine Bewertungen
- Disorders of The Psittacine Gastrointestinal TractDokument21 SeitenDisorders of The Psittacine Gastrointestinal TractRebeca GalvãoNoch keine Bewertungen
- Digestive System Overview: Parts and FunctionsDokument62 SeitenDigestive System Overview: Parts and FunctionsKeturah IbrahimNoch keine Bewertungen
- Digestive System of A RabbitDokument5 SeitenDigestive System of A RabbitWendell John Noda MirandaNoch keine Bewertungen
- NPTEL Basic Biology Digestive SystemDokument74 SeitenNPTEL Basic Biology Digestive SystemBikashGuptaNoch keine Bewertungen
- Chicken AnatomyDokument6 SeitenChicken AnatomyavduqueNoch keine Bewertungen
- Chicken Anatomy and Physiology Chapter OverviewDokument30 SeitenChicken Anatomy and Physiology Chapter OverviewJustine Mae Balodong BumalayNoch keine Bewertungen
- Lab Ex 3 CDokument7 SeitenLab Ex 3 Cs.manuel.kyrraaliahNoch keine Bewertungen
- Digestion and AbsorptionDokument6 SeitenDigestion and AbsorptionMahafuj DudhaknojNoch keine Bewertungen
- Digestive SystemDokument6 SeitenDigestive SystemAiman AfzalNoch keine Bewertungen
- gastrointestinal-physiology-and-nutrition-in-wild-birdsDokument8 Seitengastrointestinal-physiology-and-nutrition-in-wild-birdsLayna RiNoch keine Bewertungen
- Ekomorfo1 - Baru (Compatibility Mode)Dokument9 SeitenEkomorfo1 - Baru (Compatibility Mode)Nisa WidiaswaraNoch keine Bewertungen
- The Digestive SystemDokument23 SeitenThe Digestive SystemJenderal Yohanes100% (1)
- Ruminant Digestive SystemDokument12 SeitenRuminant Digestive SystemMacharia JosephNoch keine Bewertungen
- E Pathshala File 801642381308Dokument48 SeitenE Pathshala File 801642381308Aijaz AhmadNoch keine Bewertungen
- Anatomy and Physiology of The Ruminant Animal Digestive SystemDokument19 SeitenAnatomy and Physiology of The Ruminant Animal Digestive SystemNatanga GodfreyNoch keine Bewertungen
- Monogastric: Single-Chambered StomachDokument4 SeitenMonogastric: Single-Chambered StomachFarhan Rezki MNoch keine Bewertungen
- Earth and Life Module Week 16Dokument9 SeitenEarth and Life Module Week 16Austin Capal Dela CruzNoch keine Bewertungen
- 1-Equine Digestion PowerPointDokument15 Seiten1-Equine Digestion PowerPointMazhar FaridNoch keine Bewertungen
- Mca Feb2011Dokument4 SeitenMca Feb2011Agnes OctaviaNoch keine Bewertungen
- Digestive System 2021Dokument29 SeitenDigestive System 2021fashion20trendsNoch keine Bewertungen
- Koya University Faculty of Science and Health Biology DepartmentDokument4 SeitenKoya University Faculty of Science and Health Biology DepartmentRayan AliNoch keine Bewertungen
- Human Digestive System AnatomyDokument59 SeitenHuman Digestive System AnatomyTarun AroraNoch keine Bewertungen
- PRACTICA FINAL DE INGLE. DIGESTIVE SYSTEM. AnaDokument18 SeitenPRACTICA FINAL DE INGLE. DIGESTIVE SYSTEM. AnaChantal PantaleonNoch keine Bewertungen
- Anatomy DigestiveDokument8 SeitenAnatomy DigestiveGalih Kurniawan100% (2)
- Ferret NutritionDokument23 SeitenFerret NutritionlauraNoch keine Bewertungen
- Animal Nutrition and FeedingDokument29 SeitenAnimal Nutrition and FeedingMaster MenteNoch keine Bewertungen
- Ruminant Animal Nutrition GuideDokument44 SeitenRuminant Animal Nutrition GuideYusnaini Arifin100% (1)
- Rumen Physiology and AnatomyDokument6 SeitenRumen Physiology and AnatomyAgnes OctaviaNoch keine Bewertungen
- Herbivore-: Animals Have Different Ways of Digesting The Food They Eat. Herbivores Are Animals That Eat PlantsDokument4 SeitenHerbivore-: Animals Have Different Ways of Digesting The Food They Eat. Herbivores Are Animals That Eat PlantsAFIQAH ZNoch keine Bewertungen
- Vet Histology Lecture Notes on Digestive System IDokument62 SeitenVet Histology Lecture Notes on Digestive System IDEV3LLS100% (3)
- Cell is the smallest unit of lifeDokument8 SeitenCell is the smallest unit of lifeAnsel Francis RapsingNoch keine Bewertungen
- Hetrotrophic Nutrition o Level PDFDokument15 SeitenHetrotrophic Nutrition o Level PDFTimothy PetersonNoch keine Bewertungen
- Mod3 PDFDokument74 SeitenMod3 PDFHfNoch keine Bewertungen
- 1 Digestion in Teleost Fishes - 2Dokument23 Seiten1 Digestion in Teleost Fishes - 2Evi Nurul IhsanNoch keine Bewertungen
- Anatomy and Physiology of The Rabbit and Rodent GaDokument10 SeitenAnatomy and Physiology of The Rabbit and Rodent GaSabrina CalabranoNoch keine Bewertungen
- Avian Digestive System - ExtensionDokument6 SeitenAvian Digestive System - ExtensionShakil MahmodNoch keine Bewertungen
- Digestive Presentation Animal Related SystemsDokument40 SeitenDigestive Presentation Animal Related Systemsapi-293732226Noch keine Bewertungen
- Rabbit Rundown: A Data Engineers Guide To Rabbitats, Rabbitology, and Rabbitisms: GuidesVon EverandRabbit Rundown: A Data Engineers Guide To Rabbitats, Rabbitology, and Rabbitisms: GuidesNoch keine Bewertungen
- Soil Erosion RiskDokument22 SeitenSoil Erosion RiskMogie TalampasNoch keine Bewertungen
- Sony KDL-32BX320 - 32BX321 - 32BX420 - 32BX421 - 40BX421 - 46BX420 - 46BX421... AZ2-UK ChassisDokument74 SeitenSony KDL-32BX320 - 32BX321 - 32BX420 - 32BX421 - 40BX421 - 46BX420 - 46BX421... AZ2-UK Chassismartijn16340% (1)
- SchedDokument1 SeiteSchedAaron WoodardNoch keine Bewertungen
- Methods For AASDokument80 SeitenMethods For AASAaron WoodardNoch keine Bewertungen
- 04 Concept and Use of The Process Approach For Management SystemsDokument12 Seiten04 Concept and Use of The Process Approach For Management SystemsAaron WoodardNoch keine Bewertungen
- DPSIR OverviewDokument10 SeitenDPSIR OverviewAaron WoodardNoch keine Bewertungen
- Birds of PalawanDokument24 SeitenBirds of PalawanAaron WoodardNoch keine Bewertungen
- PrintDokument1 SeitePrintAaron WoodardNoch keine Bewertungen
- E CatalogueDokument47 SeitenE CatalogueDr. Bill MotendeNoch keine Bewertungen
- Nestle Catalogue 2021Dokument18 SeitenNestle Catalogue 2021T S DhanashreeNoch keine Bewertungen
- Contoh Soalan TOV Year 5Dokument8 SeitenContoh Soalan TOV Year 5Fatin Nur HafizahNoch keine Bewertungen
- Listado #15 Work and Travel 2024 - Empleadores - Nov 29 2023Dokument18 SeitenListado #15 Work and Travel 2024 - Empleadores - Nov 29 2023MiguelNoch keine Bewertungen
- Rephrasing C1Dokument7 SeitenRephrasing C1Pablo AlejandroNoch keine Bewertungen
- The PH of Beverages in The United StatesDokument9 SeitenThe PH of Beverages in The United StatesRazty BeNoch keine Bewertungen
- Arup Explores - The Future of Urban Agriculture - Final ReportDokument48 SeitenArup Explores - The Future of Urban Agriculture - Final Reportvishnu vijayanNoch keine Bewertungen
- Alcoholic BeveragesDokument101 SeitenAlcoholic BeveragesAmazing Grace Dela Cruz100% (2)
- Indigenous Fermented Food and BeveragesDokument5 SeitenIndigenous Fermented Food and BeveragesdodecadidoNoch keine Bewertungen
- LAPORAN RATA-RATA HARGA BELIDokument315 SeitenLAPORAN RATA-RATA HARGA BELIfadjaroeddinNoch keine Bewertungen
- Animals and Birds of Rainy SeasonDokument3 SeitenAnimals and Birds of Rainy SeasonMamta M RNoch keine Bewertungen
- 7 Causes of Disease-Dr Sherry RogersDokument10 Seiten7 Causes of Disease-Dr Sherry RogersJose RodriguezNoch keine Bewertungen
- IFS Trend Risk Report May 2023 1687538806Dokument12 SeitenIFS Trend Risk Report May 2023 1687538806Gisele JometónNoch keine Bewertungen
- Integrated Rice and Duck Farming - Takao Furuno CaseDokument2 SeitenIntegrated Rice and Duck Farming - Takao Furuno CaseRija Ménagé100% (1)
- BÀI TẬP TRẮC NGHIỆM IDIOMSDokument55 SeitenBÀI TẬP TRẮC NGHIỆM IDIOMSThành Long Tứ ĐỗNoch keine Bewertungen
- FDA Comprehensive Organization Chart 20170925Dokument1 SeiteFDA Comprehensive Organization Chart 20170925gabyNoch keine Bewertungen
- IILM Institute For Higher Education: Quality Management ProjectDokument10 SeitenIILM Institute For Higher Education: Quality Management ProjectPratibha JainNoch keine Bewertungen
- Anjali Shukla Research Report FileDokument55 SeitenAnjali Shukla Research Report FileAnjali ShuklaNoch keine Bewertungen
- Breakfast Vocabulary Esl Multiple Choice Test For KidsDokument2 SeitenBreakfast Vocabulary Esl Multiple Choice Test For KidsNada TrifkovicNoch keine Bewertungen
- Orange and Cinnamon MacaronsDokument2 SeitenOrange and Cinnamon MacaronscakecrumbsljNoch keine Bewertungen
- Tariff Rulings MARCH 2022 Final 3 1Dokument22 SeitenTariff Rulings MARCH 2022 Final 3 1milanNoch keine Bewertungen
- Satvic Kids BookDokument29 SeitenSatvic Kids Bookmahesh gole100% (4)
- Thirst - C.A. JohnsonDokument92 SeitenThirst - C.A. JohnsonMaddy MullaheyNoch keine Bewertungen
- Christian Dieting Programs-Like Gwen Shamblin's Weigh Down Diethelp Believers Pray Off The Pounds. But What Deeper Messages Are They Sending About Faith and Fitness?Dokument10 SeitenChristian Dieting Programs-Like Gwen Shamblin's Weigh Down Diethelp Believers Pray Off The Pounds. But What Deeper Messages Are They Sending About Faith and Fitness?laurapc55100% (1)
- R Digest Class Capers 1Dokument79 SeitenR Digest Class Capers 1sarah rasydNoch keine Bewertungen
- Amarnath JI Yatra by Helicopter: Min - 2 Pax (Nett)Dokument3 SeitenAmarnath JI Yatra by Helicopter: Min - 2 Pax (Nett)Deepak BothraNoch keine Bewertungen
- Hampshire College Student Café Co-op ProposalDokument6 SeitenHampshire College Student Café Co-op ProposalAlok ShenoyNoch keine Bewertungen
- La Maison 1888 Lunch Menu EngDokument2 SeitenLa Maison 1888 Lunch Menu Engn4ff5bdm7qNoch keine Bewertungen
- Pizza Hut ProjectDokument14 SeitenPizza Hut Projectsakshi23118679% (14)
- USBN 2018-2019 English Exam Practice QuestionsDokument10 SeitenUSBN 2018-2019 English Exam Practice QuestionsauleeahgittaNoch keine Bewertungen