Beruflich Dokumente
Kultur Dokumente
Dissertation XingyuanChen - Final
Hochgeladen von
sita_adhiOriginalbeschreibung:
Originaltitel
Copyright
Verfügbare Formate
Dieses Dokument teilen
Dokument teilen oder einbetten
Stufen Sie dieses Dokument als nützlich ein?
Sind diese Inhalte unangemessen?
Dieses Dokument meldenCopyright:
Verfügbare Formate
Dissertation XingyuanChen - Final
Hochgeladen von
sita_adhiCopyright:
Verfügbare Formate
Study of Water Dynamics in the Soil-Plant-Atmospheric Continuum in a
Water-Controlled Ecosystem
by
Xingyuan Chen
B.ENG. (Tsinghua University, Beijing, China) 2000
M.Phil. (Hong Kong University of Science and Technology) 2002
M.A. (University of California, Berkeley) 2007
A dissertation submitted in partial satisfaction of the
requirements for the degree of
Doctor of Philosophy
in
Engineering-Civil and Environmental Engineering
in the
GRADUATE DIVISION
of the
UNIVERSITY of CALIFORNIA at BERKELEY
Committee in charge:
Professor Yoram Rubin, Co-Chair
Professor Dennis D. Baldocchi, Co-Chair
Professor Mark Stacy
Professor David Brillinger
Spring 2009
The dissertation of Xingyuan Chen is approved:
Co-Chair Date
Co-Chair Date
Date
Date
University of California at Berkeley
Spring 2009
Study of Water Dynamics in the Soil-Plant-Atmospheric Continuum in a
Water-Controlled Ecosystem
Copyright Spring 2009
by
Xingyuan Chen
1
Abstract
Study of Water Dynamics in the Soil-Plant-Atmospheric Continuum in a
Water-Controlled Ecosystem
by
Xingyuan Chen
Doctor of Philosophy in Engineering-Civil and Environmental Engineering
University of California at Berkeley
Professor Yoram Rubin, Co-Chair
Professor Dennis D. Baldocchi, Co-Chair
The study of water exchange between soil, plants, and the atmosphere in response to sea-
sonal or periodic droughts is critical to modeling the hydrologic cycle and biogeochemical
processes in water-controlled ecosystems. This dissertation consists of four essential parts
of water dynamics studies in an oak Savanna ecosystem.
The rst study characterizes changes in evaporation and transpiration under water stress.
The inuence of soil moisture on evapotranspiration at the stand scale is studied using
correlations between tower-based evapotranspiration measurements and representative soil
moisture obtained by aggregating point measurements. The observed pattern of this effect
is found in agreement with an existing model that features a linear reduction of the evap-
2
otranspiration when soil moisture falls below a critical value. The model parameters are
inferred using a Bayesian framework and they are found to vary from year to year due to
climate variability. The comparison between various aggregations of soil moisture at the
stand scale from point measurements demonstrates that the spatial variability of the soil
moisture as well as the water uptake capacity limited by the root biomass need be taken
into account to produce a model that is most resistant to inter-annual variability.
The second study re-evaluated the theoretical basis of sap ow measurements using
heat ratio method, whose fundamental basis was built on an idealized solution to heat trans-
port process in sapwood. An improved solution is developed to model the same process
with more realistic assumptions. Extensive comparisons on the difference of calculated
temperature elds by idealized solution and improved solution reveal that most signicant
discrepancy occurs around the early times, whereas the difference diminishes over late time
window. This study also present changes in the fundamental equation of heat ratio method
to account for asymmetric probe alignments in practice.
The third study is on determining probe geometry and wood thermal diffusivity for
sap ow measurements using heat ratio method. A statistical framework is presented to
simultaneously estimate wood thermal diffusivity and probe geometry from in-situ heat re-
sponse curves collected by the implanted probes of heat ratio apparatus. Conditioned on the
heat response data, the parameters are inferred using a Bayesian inversion technique with
Markov chain Monte Carlo (MCMC) sampling method. This procedure not only provides
a systematic yet non-destructive to estimate the crucial parameters for sap ow calculation,
3
it also enables direct quantication of uncertainty in estimated sap ow velocity. Experi-
ments using synthetic data show that multiple tests on the same apparatus are essential to
obtain reliable and accurate solutions, and the uncertainty in posterior distributions of the
parameters is inuenced by the prior knowledge on the probe geometry or heating power.
When applied to eld conditions, multiple tests are conducted during different seasons and
automated using the existing data logging system. The seasonality of wood thermal diffu-
sivity is obtained as a by-product of the parameter estimation process, and it is affected by
both moisture content and temperature. Empirical factors are introduced to account for the
inuence of non-ideal probe geometry on the estimation of heat pulse velocity, and they
are estimated in this study as well. The proposed methodology is ready to be applied to
calibrate existing heat ratio sap ow systems at other sites. It is especially useful when
alternative transpiration calibration device such as lysimeter is not available.
The fourth study investigated how an individual plant adjusts transpiration under the
stressed conditions using continuous transpiration and soil moisture measurements. The
objective is to appropriately calculate the potential transpiration, which is often needed
in soil water dynamics models. The alternating conditional expectation (ACE) method is
implemented to identify the optimal functional dependence of bulk canopy conductance
on various environmental stresses including vapor pressure decit, net radiation, and soil
moisture. A multiplicative form of stress functions is found to be appropriate for the tree
studied. The functional form of each individual stress is determined based on the optimal
transformations identied by the ACE method, and MCMCis then implemented to estimate
4
the model parameters. The continuous transpiration and soil moisture data are also used
to investigate the water budget on the tree over the growing season of 2007. It is found
that tree transpires much more water than what is provided by the root zone soil water
during dry season, which is a strong evidence of tree tapping water from deeper soil and
groundwater in dry seasons. This factor need be accounted for in further soil dynamics
modeling by specifying appropriate boundary conditions.
Professor Yoram Rubin
Dissertation Committee Co-Chair
Professor Dennis D. Baldocchi
Dissertation Committee Co-Chair
i
To my family
and
In memory of Jianzhong Guo
ii
Contents
List of Figures v
List of Tables x
1 Introduction 1
1.1 Problem Overview . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1
1.2 Experimental Site Information . . . . . . . . . . . . . . . . . . . . . . . . 6
1.3 Chapter Organization . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9
2 Observations and Stochastic Modeling of Soil Moisture Control on Evapotran-
spiration in a Californian Oak Savanna 10
2.1 Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11
2.2 Data Acquisition . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14
2.3 Theoretical Background . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16
2.4 Results and Discussions . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
2.4.1 Micrometeorological Observations and Soil Moisture . . . . . . . . 18
2.4.2 Soil Moisture Effects on ET . . . . . . . . . . . . . . . . . . . . . 24
2.4.3 Parameterization Results and Discussions . . . . . . . . . . . . . . 31
2.4.4 Predicting ET with Calibrated Model . . . . . . . . . . . . . . . . 35
2.5 Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 37
3 Theoretical Analysis for Sap Flow Measurements using Heat Ratio Method 40
3.1 Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40
3.2 Theory . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42
3.2.1 Heat transport equation in sapwood . . . . . . . . . . . . . . . . . 42
3.2.2 Marshalls solution . . . . . . . . . . . . . . . . . . . . . . . . . . 43
3.2.3 Improved solutions . . . . . . . . . . . . . . . . . . . . . . . . . . 44
3.3 Results and Discussions . . . . . . . . . . . . . . . . . . . . . . . . . . . . 47
3.3.1 Comparison of calculated temperature elds . . . . . . . . . . . . . 47
3.3.2 Impacts on HRM . . . . . . . . . . . . . . . . . . . . . . . . . . . 55
iii
3.4 Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 56
4 On Determining Wood Thermal Diffusivity and Probe Geometry Using In-Situ
Heat Response Curves for Sap Flow Measurements 60
4.1 Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 60
4.2 Materials and Methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . 64
4.2.1 Data acquisition . . . . . . . . . . . . . . . . . . . . . . . . . . . 64
4.2.2 Parameter Estimation Method . . . . . . . . . . . . . . . . . . . . 65
4.3 Results and Discussions . . . . . . . . . . . . . . . . . . . . . . . . . . . . 67
4.3.1 Model verication . . . . . . . . . . . . . . . . . . . . . . . . . . 67
4.3.2 Applications to eld data . . . . . . . . . . . . . . . . . . . . . . . 75
4.4 Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 77
5 Monitoring and Modeling Water Dynamics at Plant Scale 81
5.1 Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 81
5.2 Materials and Methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . 84
5.2.1 Data acquisition . . . . . . . . . . . . . . . . . . . . . . . . . . . 84
5.2.2 Tree transpiration model under water stress . . . . . . . . . . . . . 89
5.3 Results and Discussions . . . . . . . . . . . . . . . . . . . . . . . . . . . . 91
5.3.1 Meteorological and water status observations . . . . . . . . . . . . 91
5.3.2 Sap ow observations . . . . . . . . . . . . . . . . . . . . . . . . 95
5.3.3 Water balance calculation . . . . . . . . . . . . . . . . . . . . . . 99
5.3.4 Parameter estimation results for bulk canopy conductance . . . . . 99
5.4 Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 110
6 Summary 112
A Parameter estimation for the Feddes Model 140
A.1 Parameter Estimation Method . . . . . . . . . . . . . . . . . . . . . . . . 140
A.1.1 Principle of Minimum Relative Entropy . . . . . . . . . . . . . . . 141
A.1.2 Markov Chain Monte Carlo Method . . . . . . . . . . . . . . . . . 142
A.1.3 Inference Using WinBUGS . . . . . . . . . . . . . . . . . . . . . 144
B Regression Relations used to Extend TDRMeasurements in 2003-2005 at Tonzi
Site 146
C Data Logger Program for Data Collection 155
D Sapwood Moisture Relations of Sap Flow Measurements 162
D.1 Moisture Dependence of Thermal Diffusivity . . . . . . . . . . . . . . . . 163
D.2 Moisture Dependence of Conversion from Heat Pulse Velocity to Sap Flux
Density . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 165
D.3 An Example . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 165
iv
E Alternating Conditional Expectation Algorithm 167
v
List of Figures
1.1 Location of experimental site overlaid on the California ecoregions map
(USEPA, 2000) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7
1.2 Schematic of TDR locations overlaid on IKONOS panchromatic imagery
with 1m resolution . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
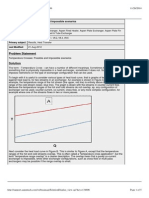
2.1 Regulation of soil moisture on ET (After Feddes, 1978; Laio et al., 2001) . 12
2.2 Meteorological conditions, soil moisture and measured ET at the Tonzi site.
The plots from top to bottom are daytime available energy, daytime air tem-
perature, daytime relative humidity and daily precipitation, depth-averaged
soil moisture (the series labeled as overall is the averaged measurements
by all TDR probes), and measured evapotranspiration . . . . . . . . . . . . 19
2.3 Regression of TDR measurements under canopy on the Theta probe mea-
surements in 2005. Theta probe measurements are daily averages calcu-
lated from continuous data on half-hour basis. TDR measurements are av-
erages calculated from TDRs located under tree canopies. The regression
equations are shown along with the regression lines. . . . . . . . . . . . . . 21
2.4 Meteorological conditions, soil moisture and measured ET at the Tonzi
site in 2003. The plots from top to bottom are daytime available energy,
daytime air temperature, daytime relative humidity and daily precipitation,
depth-averaged soil moisture (the series labeled as overall is the averaged
measurements by all TDR probes), and measured evapotranspiration . . . . 23
2.5 Time series plots of E
act
/E
eq
for (a) overstory, (b) trees and (c) understory . 25
2.6 Dependence of E
act
/E
eq
on soil moisture measured by the Theta probes . . 27
2.7 Dependence of E
act
/E
eq
on interpolated soil moisture measured by the
TDR probes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 28
2.8 Dependence of E
act
/E
eq
on root density weighted soil moisture measured
by the TDR probes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30
2.9 Posterior marginal distributions of the Feddes Model parameters . . . . . . 33
vi
2.10 Realizations of the Feddes Model compared to the data for the overstorys
ET in 2003 (Scatters are the data and lines are the realizations of the Feddes
Model) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36
2.11 Prediction of ET in 2004 and 2005 with model calibrated using data in
2003 (Scatters are the mean values of the predictions and the dashed lines
delineate the range within two standard deviations from the means) . . . . . 36
3.1 Diagram of heat ratio apparatus . . . . . . . . . . . . . . . . . . . . . . . . 41
3.2 Example of heat response curves taken downstream and upstream from the
heater probe and the corresponding time series of log-ratio of temperature
increases. The dashed line in the right panel is the log-ratio calculated from
average temperature rise over t=60-100s following the heat pulse. . . . . . 44
3.3 The temperature increases calculated from various theoretical models. Our
solution is labeled as Chens . . . . . . . . . . . . . . . . . . . . . . . . . 47
3.4 Difference in temperature rise between Marshalls solution and our solu-
tion as affected by xlocation and heat pulse velocity for L = 7 cm, =
0.0030 cm
2
/s, y = 0 cm, and z = 1.0 cm. The difference is calculated as
[
_
T
Marshall
T
Chen
_
/T
Chen
[. . . . . . . . . . . . . . . . . . . . . . . . . . 49
3.5 Difference in temperature rise between Marshalls solution and our solution
as affected by heat pulse velocity for L = 7 cm, = 0.0030 cm
2
/s, x =
0.6 cm, z = 1.0 cm, and v
h
= 30 cm/hr. The difference is calculated as
[
_
T
Marshall
T
Chen
_
/T
Chen
[. . . . . . . . . . . . . . . . . . . . . . . . . . 50
3.6 Difference in temperature rise between Marshalls solution and our solu-
tion as affected by y-coordinate for L = 7 cm, = 0.0030 cm
2
/s, x =
0.6 cm, z = 1.0 cm, and v
h
= 30 cm/hr.The difference is calculated as
[
_
T
Marshall
T
Chen
_
/T
Chen
[. . . . . . . . . . . . . . . . . . . . . . . . . . 51
3.7 Difference in temperature rise between Marshalls solution and our solu-
tion as affected by y-coordinate for L = 7 cm, = 0.0030 cm
2
/s, x =
0.6 cm, y = 0.0 cm, and v
h
= 30 cm/hr.The difference is calculated as
[
_
T
Marshall
T
Chen
_
/T
Chen
[. . . . . . . . . . . . . . . . . . . . . . . . . . 52
3.8 Difference in temperature rise between Marshalls solution and our solution
as affected by the sapwood depth for = 0.0030 cm
2
/s, x = 0.6 cm, y =
0.0 cm, z = 1.0 cm, and v
h
= 30 cm/hr. The difference is calculated as
[
_
T
Marshall
T
Chen
_
/T
Chen
[. . . . . . . . . . . . . . . . . . . . . . . . . . 53
3.9 Difference in temperature rise between Marshalls solution and our solu-
tion as affected by the sapwood depth for L = 7 cm, x = 0.6 cm, y =
0.0 cm, z = 1.0 cm, and v
h
= 30 cm/hr. The difference is calculated as
[
_
T
Marshall
T
Chen
_
/T
Chen
[. . . . . . . . . . . . . . . . . . . . . . . . . . 54
3.10 Contours of empirical factors versus spacing asymmetry in x- and y- direc-
tions. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 57
vii
4.1 Posterior distributions of parameters with known heat amount, wood den-
sity and wood specic heat capacity. The limits of x-axis represent the
bounds imposed on parameters as prior distributions. The solid vertical
lines represent the true parameter values and dashed vertical lines are the
mean values calculated from the posterior distributions. . . . . . . . . . . . 70
4.2 Model tting and statistical distribution of estimated heat pulse velocities
for the test case with known heat amount, wood density, and wood specic
heat capacity. In the left panel, each solid line represents a model t to the
data by a parameter set from the MCMC method. In the right panel, solid
vertical lines represent the true heat pulse velocities and dashed vertical
lines are the mean values calculated from the posterior distributions. . . . . 71
4.3 Posterior distributions of parameters with unknown heat amount, wood
density and wood specic heat capacity. The solid lines represent the true
parameter values and dashed vertical lines are the mean values calculated
from the posterior distributions. . . . . . . . . . . . . . . . . . . . . . . . 73
4.4 Model tting and statistical distributions of estimated heat pulse velocities
for the test case with unknown heat amount, wood density, and wood spe-
cic heat capacity. In the left panel, each solid line represents a model t
to the data by a parameter set from the MCMC method. In the right panel,
solid vertical lines represent the true heat pulse velocities and dashed ver-
tical lines are the mean values calculated from the posterior distributions. . 74
4.5 Seasonal change of inferred wood thermal diffusivity with precipitation
and wood temperature. Means of wood thermal diffusivity are shown in
stars and the vertical lines denote the uncertainty range within one standard
deviation. Wood temperature is shown in circles and precipitation is shown
in bars. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 76
4.6 Model tting to data and statistical distributions of estimated heat pulse ve-
locity for the eld test.In the left panel, each solid line represents a model
t to the data by a parameter set from the MCMC method. In the right
panel, dashed vertical lines are the mean values calculated from the poste-
rior distributions. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 78
5.1 Schematic of measurement equipments installed around each experimental
tree . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 85
5.2 Calibration curves for EC-5 sensors . . . . . . . . . . . . . . . . . . . . . 87
5.3 Meteorological and water status observations in 2007 and 2008. Soil mois-
ture data and air temperature data are averaged values during daytime (8am-
6pm). Predawn leaf water potential (LWP) were averaged from measure-
ments taken at two other trees within the footprint of the overstory eddy
covariance tower. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 93
viii
5.4 Soil water retention curve at experimental site. The range of water re-
tention curves (in solid lines) were obtained from soil texture and bulk
density using pedotransfer functions in Rosetta database (USDA, 1999),
which adopts van Genuchten model [van Genuchten, 1980]. Data points in
squares were from samples analyzed at the DANR Analytical Soils Labo-
ratory, University of California-Davis. Data shown in dots were obtained
by Liukang Xu in 2002, using a WP4 Dewpoint PotentiaMeter. . . . . . . 94
5.5 Heat pulse velocities measured from the bottom set of sap ow sensors.
Inside and outside probe pairs are 2.5 cm and 1 cm into the tree sapwood,
respectively. The scatters are mean values calculated from MCMC samples
and dashed lines are plus minus two standard deviations from the mean value. 96
5.6 Relation between tree diameter and sapwood area. Measurements of diam-
eter and sapwood area were conducted on fresh tree samples cut down by
the ranch owner occasionally. The boundary between sapwood and heart-
wood was identied from the change of color. . . . . . . . . . . . . . . . . 97
5.7 Daily transpiration measured from bottom set of sap ow sensors. The
scatters are mean values calculated from MCMC samples and dashed lines
are two standard deviations from the mean value. . . . . . . . . . . . . . . 98
5.8 Daily water balance. The range of negative change in soil water is cut to
enlarge the scale of the plot. The actual values for the two large negative
changes are -28mm and -60mm, respectively. . . . . . . . . . . . . . . . . 100
5.9 Weekly water balance. . . . . . . . . . . . . . . . . . . . . . . . . . . . . 101
5.10 Optimal transformations as determined by ACE. The multiple R
2
=0.99.
The units of G
c
and VPD are mm/s and kPa, respectively. . . . . . . . . . 103
5.11 Optimal transformation of log(G
c
) vs the sum of the optimal transforma-
tions of log(VPD), SWC, and R
n
. The solid line is the 1:1 line. . . . . . . . 104
5.12 Posterior marginal distributions of the G
c
stress function parameters. The
vertical dashed lines are the mean values calculated from the posterior sam-
ples. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 106
5.13 Model tting to observed transpiration and bulk canopy conductance. The
solid lines are 1:1 line . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 107
5.14 Diagnosis plots of residuals versus dependent variables. . . . . . . . . . . . 108
5.15 Potential transpiration calculated in 2007 compared to measured actual
transpiration . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 109
B.1 Regression of average TDR measurements on the Theta probe measure-
ments in 2003. Theta probe measurements are daily averages calculated
from continuous data on half-hour basis. TDR measurements are averages
calculated from all TDRs located in the site. The regression equations are
shown along with the regression lines. . . . . . . . . . . . . . . . . . . . . 147
ix
B.2 Regression of TDR measurements under tree canopies on the Theta probe
measurements in 2003. Theta probe measurements are daily averages cal-
culated from continuous data on half-hour basis. TDR measurements are
averages calculated fromTDRs located under tree canopies. The regression
equations are shown along with the regression lines. . . . . . . . . . . . . . 148
B.3 Regression of TDR measurements in open spaces on the Theta probe mea-
surements in 2003. Theta probe measurements are daily averages calcu-
lated from continuous data on half-hour basis. TDR measurements are
averages calculated from TDRs located in the open spaces (not covered by
tree canopy). The regression equations are shown along with the regression
lines. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 149
B.4 Regression of average TDR measurements on the Theta probe measure-
ments in 2004. Theta probe measurements are daily averages calculated
from continuous data on half-hour basis. TDR measurements are averages
calculated from all TDRs located in the site. The regression equations are
shown along with the regression lines. . . . . . . . . . . . . . . . . . . . . 150
B.5 Regression of TDR measurements under tree canopies on the Theta probe
measurements in 2004. Theta probe measurements are daily averages cal-
culated from continuous data on half-hour basis. TDR measurements are
averages calculated fromTDRs located under tree canopies. The regression
equations are shown along with the regression lines. . . . . . . . . . . . . . 151
B.6 Regression of TDR measurements in open spaces on the Theta probe mea-
surements in 2004. Theta probe measurements are daily averages calcu-
lated from continuous data on half-hour basis. TDR measurements are
averages calculated from TDRs located in the open spaces (not covered by
tree canopy). The regression equations are shown along with the regression
lines. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 152
B.7 Regression of average TDR measurements on the Theta probe measure-
ments in 2005. Theta probe measurements are daily averages calculated
from continuous data on half-hour basis. TDR measurements are averages
calculated from all TDRs located in the site. The regression equations are
shown along with the regression lines. . . . . . . . . . . . . . . . . . . . . 153
B.8 Regression of TDR measurements in open spaces on the Theta probe mea-
surements in 2005. Theta probe measurements are daily averages calcu-
lated from continuous data on half-hour basis. TDR measurements are
averages calculated from TDRs located in the open spaces (not covered by
tree canopy). The regression equations are shown along with the regression
lines. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 154
D.1 Dependence of wood thermal diffusivity and conversion factor on wood
moisture content . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 166
x
List of Tables
4.1 Parameters estimated by MCMC method with known probe geometry com-
pared to true values . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 73
D.1 Denitions of some concepts related to wood moisture content . . . . . . . 163
xi
Acknowledgements
First and foremost, I wish to express my sincere gratitude to my advisors, Professor
Yoram Rubin and Professor Dennis D. Baldocchi. They not only introduced me to the eld
of ecohydrology, but also have provided me with continuous guidance and encouragement.
I have benetted enormously from their expertise and enthusiasm in their distinctive elds.
The discussions with them during my PHD study have inspired many great ideas, based on
which I was able to win the Horton Research Grant from the American Geophysical Union
to conduct eld works. They have been providing invaluable feedbacks and assistances to
my research and dissertation writing in time. I also thank them for always pushing me to
think deep about why we are doing the work and to set clear goals. Without their support
and encouragement, this dissertation would not have existed.
I would also like to thank Professor Mark Stacy and Professor David Brillinger for
being on my dissertation committee. They have given me many valuable comments and
constructive suggestions during our communications.
I would like to thank Ted Hehn for his numerous help on manufacturing sap ow probes
and setting up the experiments in eld. I also thank Mr. Russell Tonzi for providing access
to his ranch for scientic research.
My thanks are due to all my colleagues, Zhangshuan Hou, Felipe Barrows, Gretchen R.
Miller, Siyan Ma, Qi Chen, Jessica Osuna, and Youngryel Ryu, for their help and collabo-
ration in data collection, and for many inspiring discussions.
I am deeply indebted to my dear friends over the years, Jianzhong He, Lili Ji, Shaonan
xii
Wu, Qing Ji, Ran Ding, Yang Feng, Maoyi Huang, Gang Wang, Dengfeng Sun, Qingfang
Wu, Peihua Jing, and many others. My PHD journey at Cal has been more enjoyable with
their companion.
A special Thank you belongs to my late friend, Jianzhong Guo. He was always there
when I needed help, just like a big brother, even when he was ghting against cancer. His
friendship and courage is my life-long treasure. Although he didnt see me nish my PHD
study, I know he would feel happy for me.
Last but not least, I am deeply grateful to my family for their endless love and uncon-
ditional support. It is them who made me the person who I am today and I would like to
dedicate my dissertation work to them.
1
Chapter 1
Introduction
1.1 Problem Overview
In water-limited ecosystems, root zone soil water availability becomes a controlling
factor on water, carbon and nutrient cycles [Rodriguez-Iturbe et al., 2001b; Williams and
Albertson, 2004], and they are thus called water-controlled ecosystems. The study of water
dynamics in such ecosystems is not only critical for global hydrological cycle and climate
modeling, but also for assessing ecosystem functioning in response to seasonal or periodic
droughts [Knapp et al., 2002; Porporato et al., 2004]. Such study is increasingly becoming
more important as recent climate studies suggest that periodic droughts are likely to occur
more frequently, due to reduced rainfall frequency resulting from global warming [Bell
et al., 2004; Esterling et al., 2000]. In western America, the threat of droughts could be
even worse due to loss of mountain snowpack in projected future climate [Service, 2004].
2
Study of water dynamics in water-controlled ecosystems requires an integrated ap-
proach that links biometeorology, biophysics, plant physiology and hydrology. The dif-
culties in such studies arise frominsufcient understanding of the nonlinear interactions be-
tween the various processes and their scale-dependence, and from ignoring multi-scale in-
teractions. Systematic multi-scale studies of water exchange in the soil-plant-atmospheric
continuum are still at very early stages, and require coordinated efforts of modeling de-
velopment and data acquisition at consistent scales [National Research Council (NRC),
2008]. New technology innovations from nano-materials to airborne and spaceborne re-
mote sensing sensors has made more variables accessible to measurements over a large
range of temporal-spatial scales [National Research Council (NRC), 2008]. Furthermore,
the emerging monitoring networks, such as FLUXNET [Baldocchi et al., 2001], have pro-
vided unique platforms for long-term multidisciplinary multi-scale study.
The purpose of this dissertation work is to develop a better understanding of water
transport in the soil-plant-atmospheric continuum under drought conditions, at scales rang-
ing from that of a plant (a few m
2
) to that of a stand (a few hundreds to a few thousands m
2
).
The primary focus is to investigate the regulation of plant water use by water decits and
climatic conditions and its scale dependence. Models of water stress function are exam-
ined and calibrated at both plant scale and stand scale using water exchange data acquired
at both scales.
The data necessary for model calibration include the spatial and temporal measurements
of soil water content and water uxes throughout the soil-plant-atmospheric continuum, as
3
well as climate driving forces such as net radiation, humidity and air temperature, etc. The
data acquisition efforts will focus on a Mediterranean oak-savanna ecosystem, and it will be
introduced in details in the next section. The site experiences dry and hot summer annually,
which makes it ideal for water dynamics study in water-controlled ecosystems.
Data have been collected at the site since 2001, including meteorological variables
(e.g., solar radiation, air temperature, wind speed, etc), biophysical variables (e.g., leaf area
index, leaf water potential, etc), soil moisture contents, isotopes in water and CO
2
sources,
as well as overstory and understory eddy uxes of water vapor and CO
2
. In addition to
this on-going effort, and in support of my research goals at individual plant scale, plant-
scale measurements were installed in eight trees across the footprint of the overstory eddy
covariance tower. The specic measurements on an individual tree include multiple soil
moisture probes to monitor soil moisture prole vertically and laterally, soil temperature
probes for the purpose of energy balance and soil respiration calculations, and sap ow
sensors at different tree heights for measuring tree transpiration. More detailed descriptions
on the data collection activity are available in individual chapters that utilize specic data.
While the water vapor uxes measured by the eddy covariance tower provides an es-
timation of water used by all the trees within the tower footprint, individual plant water
use is directly measured with sap ow sensors using heat-pulse technique. The heat pulse
technique uses a heat pulse as tracer and deduce the travel speed of water in porous me-
dia through the speed at which heat is transported. The heat pulse technique is known to
possess advantages of low power consumption, simple instrumentation and automated data
4
collection [Green et al., 2003]. Among all the variations of the heat pulse technique, the
heat ratio method (HRM) introduced by Burgess et al. [1998] is chosen in this study due to
its capability of detecting reverse and low ow rates and its straightforward instrumentation
[Burgess et al., 2001].
The theoretical basis underlying HRM is an analytical solution to the heat transport
equation in tree sapwood derived by Marshall [1958]. The solution, however, was re-
lied on several idealized assumptions, which are routinely violated in real applications of
HRM. Therefore, an improved solution is developed to describe the heat transport process
in sapwood with more realistic assumptions and modications to the original fundamental
equation of HRM is provided to account for the departure of eld experimental setup from
the ideal setup.
Furthermore, the successful application of HRM depends on obtaining reliable estima-
tion of sapwood thermal diffusivity and probe geometry. The empirical estimation of wood
thermal diffusivity requires information on wood moisture content and wood bulk density,
which are usually obtained through intrusive core sampling. Exact spacing and geometry
of the HRM apparatus cannot be guaranteed during installation due to the nature of the
wood matrix, especially in hardwood species such as oak and maple. To assure the quality
of the sap ow measurements acquired from the experimental site, this dissertation study
also provides a systematic, non-destructive, and replicable methodology to determine wood
thermal diffusivity and probe geometry for the sap ow measurements using the heat ratio
apparatus. The estimation of these parameters is conditioned on the time series of tem-
5
perature increases, i.e., temperature response curves, monitored by the downstream and
upstream temperature probes after a heat pulse is released by the central heating probe.
The primary advantage of the methodology is that it relies on the information that can be
obtained using the installed probes without any further disturbance to the tree.
The availability of new observations can improve our understanding of the complex
mechanisms related to water dynamics and thus enables newer model development. On
the other hand, it provides valuable information to model calibration. Successful model
calibration based on appropriate eld data through inverse modeling may prove to be an
effective tool in studies of plant water uptake [Hopmans and Bristow, 2002; Green et al.,
2006]. One challenge in model calibration is to deal with uncertainties arising from various
sources. A Bayesian approach is often chosen in model calibration for its strengths in
dealing with uncertainties arising from various sources and in introducing assumptions and
prior knowledge of the parameters through prior distributions. Another unique advantage of
Bayesian approach is the availability of many powerful inference techniques [Gelman et al.,
1995], such as Markov chain Monte Carlo (MCMC) method, which are not available to
traditional statistical approaches. The Bayesian framework with MCMC sampling methods
have been successfully implemented in many areas of studies [Clark, 2005; Clark and
Gelfand, 2006; Crainiceanu et al., 2003; Reis and Stedinger, 2005; Smith and Marshall,
2008]. Therefore, it is adopted for model calibration in this dissertation.
6
1.2 Experimental Site Information
All the eld experiments involved in this dissertation were conducted at Tonzi Ranch,
which is an active FLUXNET site located on the lower foothill of the Sierra Nevada Moun-
tains (see Figure 1.1), near Ione, California (latitude: 38.4311
N; longitude: 120.966
W;
altitude: 177 m). The site is an oak savanna woodland with scatterings of grey pine trees.
According to the meteorological records, the mean air temperature of the region is 16.6
C
and the mean annual precipitation is about 559mm, most of which occurs between October
and May [Baldocchi et al., 2004; Ma et al., 2007]. The uneven seasonal distribution of
precipitation leads to a wet and cold winter and a dry and hot summer at the study site.
The ecosystem at the site is comprised of a nonuniform distribution of trees and grasses.
The oak trees covers approximately 43% of the land at a scale up to 1km1km around the
tall tower [Kim et al., 2006] based on a high-resolution IKONOS image as shown in Figure
1.2. A recent study based on LIDAR data nds the canopy coverage to be 47% within a
200m200marea around the tower and the oak trees have heights of 9.34.3 meters [Chen
et al., 2008a]. The oak trees leaf out in the spring (around day 90) and rapidly reach full
photosynthetic potential until they senesce in the late autumn [Xu and Baldocchi, 2003].
The grasses, on the other hand, are active from late autumn to early summer and senesce
when soil water is depleted during the prolonged dry summer. The trees and grasses are
out of phase in their growing seasons with only a short overlap from April to early June.
Their different life cycles reect various mechanisms to survive through seasonal droughts
in co-existing vegetation types.
7
1. Coast Range
4. Cascades
5. Sierra Nevada
6. Chaparral/Oak Woodlands
7. Central California Valley
8. Southern California Mountains
9. Eastern Cascade Slopes and Foothills
13. Central Basin and Range
14. Mojave Basin and Range
78. Klamath Mountains
80. Northern Basin and Range
81.SonoranBasin and Range
1. Coast Range
4. Cascades
5. Sierra Nevada
6. Chaparral/Oak Woodlands
7. Central California Valley
8. Southern California Mountains
9. Eastern Cascade Slopes and Foothills
13. Central Basin and Range
14. Mojave Basin and Range
78. Klamath Mountains
80. Northern Basin and Range
81.SonoranBasin and Range
Tonzi
Ran ch
Figure 1.1: Location of experimental site overlaid on the California ecoregions map
(USEPA, 2000)
8
UTM Easting (m)
U
T
M
N
o
r
t
h
i
n
g
(
m
)
677350 677400 677450 677500 677550 677600 677650 677700
4255400
4255450
4255500
4255550
4255600
4255650
4255700
4255750
N
TDR
Understory Tower
Overstory Tower
Figure 1.2: Schematic of TDR locations overlaid on IKONOS panchromatic imagery with
1m resolution
9
The soil at Tonzi is an Auburn very rocky silt loam [Sketchley, 1965]. Our survey at
49 locations over a 200 m 200 m area found that the soil is composed of 49 pm 3%
sand, 36 2% silt and 15 1% clay (soil samples analyzed at the Division of Agriculture
and Natural Resources Analytical Soils Laboratory, University of California-Davis). Well
logs reveal that the soil layer is about 1.0 m thick and is overlaid on fractured greenstone
bedrock.
1.3 Chapter Organization
This dissertation is organized in the following way: Chapter 2 presents observations
and stochastic modeling of soil moisture control on evapotranspiration in the experimen-
tal site. This study uses data collected at stand scale and discusses the controlling role of
soil moisture on evapotranspiration from trees, grasses and the entire community. Chap-
ter 3 revisits the theoretical basis of sap ow measurements using HRM and revises the
fundamental equation of HRM to account for departure of eld conditions from idealized
ones. Chapter 4 presents a statistical technique to estimate key parameters for estimat-
ing tree transpiration from the sap ow measurements. This study is conditioned on time
series data of temperature rise responding to a heat pulse. Chapter 5 studies response of
whole-plant transpiration to soil and atmospheric water decit using data collected on an
individual tree. A summary is provided in Chapter 6.
10
Chapter 2
Observations and Stochastic Modeling
of Soil Moisture Control on
Evapotranspiration in a Californian Oak
Savanna
1
1
This chapter is based on a published article in Water Resources Research, Vol 44, W08409, doi:
10.1029/2007WR006646
11
2.1 Introduction
Oak savanna ecosystems in California are subject to soil water stress during the pro-
longed dry summer seasons. In such water-limited ecosystems, root zone soil water avail-
ability becomes a controlling factor on water, carbon and nutrient cycles [Rodriguez-Iturbe
et al., 2001b; Williams and Albertson, 2004]. Therefore, characterization of changes in
evaporation and transpiration under water stress is essential to understand the dynamics in
the soil-plant-atmosphere continuum [Feddes et al., 2001; Feddes and Raats, 2004; Teuling
et al., 2006]. Eventually these processes will contribute to the studies of the hydrological
cycle including the computation of water budgets [Milly, 1993; Botter et al., 2007] and
to the studies of biogeochemical processes in similar ecosystems [Porporato et al., 2003;
DOdorico et al., 2003].
The impacts of soil water stress on water and carbon exchange processes have been
studied experimentally and have been incorporated into various modeling efforts [Baldoc-
chi et al., 2004; Dewar, 2002; Feddes et al., 1978; Katul et al., 2003; Rodriguez-Iturbe
et al., 2001b; Tuzet et al., 2003; Williams and Albertson, 2004, 2005]. A piecewise linear
function of the dependence of evaporation and transpiration (ET) on soil moisture, shown in
Figure 2.1, is especially appealing for its simplicity and its applicability in a range of plant
functional types at leaf and stand scales [Feddes et al., 1978; Federer, 1979; Gollan et al.,
1985; Paruelo and Sala, 1995; Spittlehouse and Black, 1981]. This model will be referred
to as Feddes Model in the subsequent discussion. The Feddes Model has been adopted
in many studies as an a priori assumption with the parameters selected from literature
12
ET
1
ET
max
Stressed
Figure 2.1: Regulation of soil moisture on ET (After Feddes, 1978; Laio et al., 2001)
[Guswa et al., 2002; Laio et al., 2001; Miller et al., 2007] due to the lack of concomitant
observations of ET and soil moisture at matching scales.
Although the Feddes Model has been found efcient in many cases, the actual ET be-
havior could be different and more complex [Lagergren and Lindroth, 2002] due to the non-
linear interactions between the soil, plants and the atmosphere and their scale-dependence.
This behavior is further complicated in savannas by the coexistence of woody and herba-
ceous species, by their open heterogeneous canopies and their competition for the limited
water resources [Scholes and Archer, 1997]. All these are elements which are not in line
with the underlying assumptions of the Feddes Model.
There is only a limited body of research on the effects of soil moisture on ET in sa-
13
vannas [Detto et al., 2006; Williams and Albertson, 2004]. Therefore, the objective of our
study is to examine experimentally the applicability of the Feddes Model in a Californian
oak savanna for modeling the effect of water stress on ET of single functional types and of
their mixture at the stand scale, and to gain, through this investigation, a broader perspec-
tive on the myriad of related issues. Specic questions that we will address include: (1) Is
the Feddes Model applicable to oak savannas with heterogeneous open canopies? This is
an important question in the context of modeling water dynamics in savanna ecosystems
because ET is a crucial component in water cycling and its change under water stress needs
be taken into account in modeling; (2) How do trees and grasses in the oak savanna respond
differently to the water stress? Heterogeneity in soil moisture in savanna is both cause and
consequence of tree-grass coexistence [Rodriguez-Iturbe, 2000]. To capture the effects of
soil moistures spatial variability in water dynamics modeling, it is essential to investigate
how trees and grasses, as individual functional types, use water under seasonal droughts;
(3) Is the Feddes Model stationary in time? This is an important issue when the model char-
acterizing soil moistures control on ET is to be used in water dynamics modeling, because
large inter-annual variability of parameters would deteriorate the predictive capabilities of
models validated using historic records of undetermined duration. This specic study has
not been possible until recently, when multi-year data from the FLUXNET sites became
available; (4) What soil moisture data is needed for modeling water stress at the stand scale
and what are the alternatives for aggregating point measurements of soil moisture to the
spatial averages at the stand scale? The answer to this question will be useful for guiding
14
the acquisition of soil moisture eld data needed for model validation and prediction.
To address these questions, the observed actual ET from eddy covariance towers will
be evaluated as a function of soil moisture and potential ET. The partition of ET between
woody and herbaceous species is allowed through the deployment of overstory and under-
story eddy ux towers. A Bayesian framework with Markov chain Monte Carlo (MCMC)
method that has been applied successfully in many elds [Clark, 2005; Clark and Gelfand,
2006; Crainiceanu et al., 2003; Reis and Stedinger, 2005] is adopted to parameterize the
model that describes controlling effect of soil moisture on ET, taking into account the un-
certainties in model parameters and in eld measurements. The inter-annual variability
in the dependence of ET on soil moisture for heterogeneous environment as well as its
components is analyzed using multi-year data. The parameterized model is used for pre-
dicting the actual ET with uncertainty estimates determined using the joint distribution of
the parameters derived from the Bayesian framework.
2.2 Data Acquisition
Abroad suite of data types have been collected at the Tonzi Ranch since 2001, including
meteorological variables (e.g., solar radiation, air temperature, wind speed, precipitation,
etc), biophysical variables (e.g., leaf area index, leaf water potential, etc), soil moisture,
as well as overstory and understory eddy uxes of water vapor and CO
2
[Baldocchi et al.,
2004; Xu and Baldocchi, 2003]. The meteorological variables are needed for estimating
15
reference ET assuming sufcient water supply, which is called potential ET. The eddy
covariance measurements of water vapor ux were taken through a 23m-high overstory
tower and are viewed as representing the ET from the mixture of trees and grasses within its
300m300m fetch. In addition, eddy covariance measurements from a 2m-high understory
tower were also taken, and are viewed as representing the ET from the understory grasses
and soil surface. The difference in water vapor uxes between the overstory and understory
towers is a measure of transpiration by trees. All the measurements by the two towers were
averaged at half an hour basis.
Soil moisture at the site has been monitored using both segmented time-domain reec-
tometry (TDR) probes (Moisture Point, model 917, Environmental Sensors, Inc., Victoria,
British Columbia) and Theta probes (Delta-T Devices, model ML2-X, Cambridge, UK).
These technologies are described in detail in Blonquist et al. [2005]. The TDR probes were
installed at nine locations as shown in Figure 1.2, and each probe measures the average
volumetric soil water content at depths between 0-15, 15-30, 30-45 and 45-60cm. Among
the nine TDR probes, ve were installed under tree canopies and four in the open spaces.
TDR measurements were taken weekly or bi-weekly during eld visits. Theta probes were
installed under a tree near the tall eddy ux tower at depths of 5, 20 and 50cm, and contin-
uous soil moisture measurements were taken at half-hour interval.
16
2.3 Theoretical Background
To describe the degree to which the actual ET in the ecosystem is limited by soil wa-
ter availability, ET measured by the eddy covariance tower is compared to a reference ET
under sufcient water supply, i.e., potential ET. The latter is approximated by the Priestley-
Taylor (P-T) equation [Priestley and Taylor, 1972] as shown in the following equation,
E
p
= E
eq
=
1
s(R
n
G)
s +
, (2.1)
where E
p
is the potential ET, E
eq
is the equilibrium ET, is a scaling factor that depends
on vegetation type and climatic conditions, R
n
is the net radiation, G is the ground heat
ux density, is the latent heat of vaporization of water, s is the derivative of the saturated
vapor pressure against temperature and is the psychrometric constant.
The actual ET is assumed to be a fraction of the potential ET due to water decit
[de Bruin, 1983; Laio et al., 2001; Mahfouf et al., 1996; Rana and Katerji, 2000; Williams
and Albertson, 2004], i.e.,
E
act
= () E
p
, (2.2)
where () is the reduction coefcient of ET as a function of soil moisture. The form of
() is not assumed as prior knowledge but will be examined and compared to the Feddes
17
Model using eld data in our study. The analytical form of the Feddes Model is
() = min
_
1, max
_
0,
1
__
(2.3)
where
1
is normally referred to as permanent wilting point at which plants stop ET and
2
is the critical soil moisture value below which ET is limited by available soil moisture.
The P-T coefcient, of Eq. (2.1), is not directly measurable. The value of is
often taken as 1.26 for a wet surface [Priestley and Taylor, 1972]. However, the actual
value could deviate signicantly from 1.26 depending on the canopy structures and atmo-
spheric conditions [Baldocchi and Meyers, 1998; de Bruin and Keijman, 1979; Hikaru,
2005; Jones, 1992; McNaughton and Spriggs, 1986]. Therefore, is considered as an un-
known constant in our study, and the form of () will be evaluated through a surrogate
variable, the ratio between the actual ET and the equilibrium ET, dened as
R() =
E
act
E
eq
= (). (2.4)
R() is clearly a rescaled form of () with being the value of R() under no soil water
stress. All the variables required to determine R() are directly measured by the eddy
covariance towers. It is thus natural to extract the form of () from the observations of
R().
To examine the form of (), it is also necessary to determine whether a representative
18
soil moisture exists at the stand scale that can be used for a stand-scale Feddes Model, and
to estimate it from point measurements. The details are discussed in next section.
Once the general form of () is determined from observations, the next step is to
identify the model parameters. In most studies, the parameters that describe the effect of
soil moisture on ET are viewed as deterministic values, although in most cases they are
adopted from the past work on similar ecosystems or derived from limited eld data. Un-
certainties could arise from many sources in addition to their inherent variability, such as
measurement errors, model errors from incomplete understanding of the underlying mech-
anism, and disparity in measurement scales and modeling scales. A probabilistic approach
based on Bayesian concepts is adopted in this study to learn model parameters considering
various uncertainties. A detailed description of the method can be found in the Appendix
A.
2.4 Results and Discussions
2.4.1 Micrometeorological Observations and Soil Moisture
The data acquired between 2003 and 2005 were employed in this study. The rst three
plots in Figure 2.2 present time series of averaged daytime (8am-6pm) available energy
(R
n
G), air temperature, relative humidity and daily precipitation, all of which show
strong seasonal patterns. The summer is characterized by high temperature and available
energy, low humidity with essentially no precipitation.
19
0 50 100 150 200 250 300 365
10
0
10
20
R
n
G
(
M
J
/
m
2
)
0 50 100 150 200 250 300 365
0
10
20
30
40
A
i
r
t
e
m
p
e
r
a
t
u
r
e
(
o
C
)
0 365
0
0.5
1
1.5
R
e
l
a
t
i
v
e
h
u
m
i
d
i
t
y
0
50
100
150
P
r
e
c
i
p
i
t
a
t
i
o
n
(
m
m
/
d
a
y
)
RH Precipitation
0 50 100 150 200 250 300 365
0
0.1
0.2
0.3
0.4
S
o
i
l
m
o
i
s
t
u
r
e
(
m
3
/
m
3
)
Overall
Understory
Open space
0 50 100 150 200 250 300 365
0
0.2
0.4
0.6
Day of year in 2003
E
T
(
c
m
/
d
a
y
)
Overstory
Tree
Understory
Figure 2.2: Meteorological conditions, soil moisture and measured ET at the Tonzi site.
The plots from top to bottom are daytime available energy, daytime air temperature,
daytime relative humidity and daily precipitation, depth-averaged soil moisture (the se-
ries labeled as overall is the averaged measurements by all TDR probes), and measured
evapotranspiration
20
Two types of soil moisture data are available at the site: periodic TDR measurements
distributed within the footprint of eddy covariance tower and continuous Theta probe mea-
surements at a point. Recognizing their difference in temporal and spatial resolutions, the
spatial averages of TDR measurements in a depth segment were regressed on the daily av-
erages of the Theta probe measurements within the same depth range, using a robust linear
approach. The resulting regression lines as shown in Figure 2.3 were used to interpolate
the periodic TDR measurements to continuous ones. More regression results for years from
2003 to 2005 can be found in Appendix B. The interpolation was carried out independently
for spatial averages of all TDR measurements, for those of measurements taken under tree
canopies and for those taken in the open spaces. The overall averages of soil moisture mea-
surements represent the soil water available to the entire ecosystem, while the averaged
soil moisture under tree canopies and those in the open spaces are considered as the soil
water available to trees and grasses, respectively. Although some tree rooting systems tend
to extend beyond the edges of canopy [Casper et al., 2003] and can deplete water from
the intercanopy areas [Lefever and Lejeune, 1997; Yokozawa et al., 1998; Lejeune et al.,
1999; Rietkerk et al., 2004], there does not seem to be extensive lateral exploration of roots
beyond the canopy edge at our site based on the soil respiration study [Tang and Baldocchi,
2005]. The soil moisture measurements at different depths for the entire area, understory
area and open spaces from 2003 to 2005 are averaged vertically and are given in the third
plot of Figure 2.2, which demonstrates contrasting wet and dry conditions in winter and
summer. The interpolated average soil moisture values are considered to better capture
21
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
015cm: average of TDRs under canopy
y=0.00+0.86x
data
regression
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.03+0.93x
1530cm: average of TDRs under canopy
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.01+0.94x
3045cm: average of TDRs under canopy
Theta probe measurements
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.06+1.13x
4560cm: average of TDRs under canopy
Theta probe measurements
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
Figure 2.3: Regression of TDR measurements under canopy on the Theta probe measure-
ments in 2005. Theta probe measurements are daily averages calculated from continuous
data on half-hour basis. TDR measurements are averages calculated from TDRs located
under tree canopies. The regression equations are shown along with the regression lines.
the spatial and temporal variability at the stand scale, and this will be demonstrated in the
further analysis of this study.
Figure 2.2 also presents the observed ET time series for trees, the understory layer and
22
the overstory layer within the footprint of the overstory eddy covariance tower. The un-
derstorys ET includes ET from the grasses under tree canopies and in the open spaces, as
well as evaporation from the soil surface, whereas the overstorys ET contains ET from the
understory layer as well as ET from the trees. Although the fetch of the understory eddy
covariance tower is smaller than that of the overstory tower [Baldocchi, 1997], the obser-
vations by the understory tower are assumed to be representative of the entire understory
layer within the fetch of the overstory tower. The ET of trees is higher than the overstorys
ET because it was calculated as the difference between the overstorys ET and understorys
ET, normalized by tree canopy coverage (taken as 40%).
The plots are enlarged for 2003 in Figure 2.4 to better demonstrate their seasonal pat-
terns within the year. The seasonality of ET is controlled by the available soil moisture,
plant physiology and the atmospheric demand [Baldocchi and Xu, 2007]. It can be ob-
served from Figure 2.4 that the ET from the ecosystem follows an upward trend as the
atmospheric demand increases until the soil water supply can no longer meet the demand,
which is the point when soil moisture starts to limit ET. The understorys ET stops earlier
than the trees ET due to the shallow root prole of grasses. The trees ET, on the other
hand, is observed to continue while the soil moisture at the top 60cm barely changes, which
indicates that trees may tap deeper water sources [e.g., Lewis and Burgy, 1964]. We also
found in Figures 2.2 and 2.4 that scattered summer rains trigger pulses in ET measured by
both towers without visible change in soil moisture, which implies that summer rains are
mostly intercepted by the tree or grass canopies and are evaporated to the atmosphere.
23
0 50 100 150 200 250 300 365
10
0
10
20
R
n
G
(
M
J
/
m
2
)
0 50 100 150 200 250 300 365
0
10
20
30
40
A
i
r
t
e
m
p
e
r
a
t
u
r
e
(
o
C
)
0 365
0
0.5
1
1.5
R
e
l
a
t
i
v
e
h
u
m
i
d
i
t
y
0
50
100
150
P
r
e
c
i
p
i
t
a
t
i
o
n
(
m
m
/
d
a
y
)
RH Precipitation
0 50 100 150 200 250 300 365
0
0.1
0.2
0.3
0.4
S
o
i
l
m
o
i
s
t
u
r
e
(
m
3
/
m
3
)
Overall
Understory
Open space
0 50 100 150 200 250 300 365
0
0.2
0.4
0.6
Day of year in 2003
E
T
(
c
m
/
d
a
y
)
Overstory
Tree
Understory
Figure 2.4: Meteorological conditions, soil moisture and measured ET at the Tonzi site in
2003. The plots from top to bottom are daytime available energy, daytime air tempera-
ture, daytime relative humidity and daily precipitation, depth-averaged soil moisture (the
series labeled as overall is the averaged measurements by all TDR probes), and measured
evapotranspiration
24
2.4.2 Soil Moisture Effects on ET
Calculated E
act
/E
eq
The ratio, R() = E
act
_
E
eq
, was calculated from the meteorological and ET observa-
tions following equations (2.1), (2.2) and (2.4) for the understory layer, for the overstory
layer and for the trees, and the corresponding results are provided in Figure 2.5. Only
daytime ratios were taken into the analysis, excluding rainy and cloudy days that are not
covered by the potential ET model or by the eddy covariance technology. The small val-
ues of E
act
_
E
eq
for the trees and for the overstory layer in winter and early spring are
primarily due to the underdevelopment of tree foliage that are not taken into account in
the Priestley-Taylor equation to determine the potential ET and the insufcient available
energy because of the low solar radiation. Therefore, the estimation of () used only the
observations between the last spring rain and the rst winter rain in the same year, during
which soil moisture can be considered as the dominating limiting factor. These periods are
highlighted in Figure 2.5 by the double arrows on top of the scatters.
Inuence of representative soil moisture
Soil moisture at the site varies horizontally between canopy and inter-canopy patches
as a result of the open nature of the canopy. It is recognized that trees and grasses use
different amount of water under the same atmospheric condition and trees obtain moisture
from deeper soil storage than grasses do. These two facts lead to the vertical heterogeneity
of soil moisture in addition to its horizontal heterogeneity. To obtain a representative soil
25
0 100 200 300 400 500 600 700 800 900 1000 1100
0
0.5
1.0
1.5
2.0
E
a
c
t
/
E
e
q
(a) Overstory
0 100 200 300 400 500 600 700 800 900 1000 1100
0
0.5
1.0
1.5
2.0
E
a
c
t
/
E
e
q
(b) Trees
0 100 200 300 400 500 600 700 800 900 1000 1100
0
0.5
1.0
1.5
2.0
Day of year (20032005)
E
a
c
t
/
E
e
q
(c) Understory
Figure 2.5: Time series plots of E
act
/E
eq
for (a) overstory, (b) trees and (c) understory
26
moisture value at the stand scale from the point measurements, the aggregation strategies
in horizontal and vertical directions have impacts on accounting for contributions of point
soil moisture to the overall stand-scale soil moisture. We examined in this study various
averaging techniques to compute the representative soil moisture values as well as their
effectiveness in describing soil moisture control on stand-scale ET.
We rst assumed that only the soil moisture data measured by the Theta probes were
available, and the arithmetic averages of measurements at three depths were adopted as
the representative soil moisture values. The plots of E
act
_
E
eq
against this representative
soil moisture as shown in Figure 2.6 display considerable variations from year to year, yet
with each year following a pattern remarkably similar to the Feddes Model. Some large
values of E
act
_
E
eq
under very dry soil moisture conditions are due to scattered summer
rainfall events, which were intercepted by the tree and grass canopies and evaporated into
the atmosphere soon after.
Secondly, the representative soil moisture was taken as the vertical arithmetic average
of spatially distributed TDR measurements. The dependence of E
act
_
E
eq
on this represen-
tative soil moisture is depicted in Figure 2.7, which shows much less inter-annual variability
compared to Figure 2.6, especially for trees. The trend of each single year again follows
similar pattern as the Feddes Model.
It is observed in Figure 2.7 that in the understory layer the dependence of E
act
_
E
eq
on
soil moisture in 2005 deviates considerably from those in 2003 and 2004. This discrepancy
is possibly caused by the unusually large amount of precipitation occurring prior to the
27
0
0.2
0.4
0.6
0.8
1.0
E
a
c
t
/
E
e
q
(a) Overstory
0
0.5
1.0
1.5
2.0
E
a
c
t
/
E
e
q
(b) Trees
0.05 0.1 0.15 0.2 0.25 0.3 0.35 0.4
0
0.5
1.0
1.5
E
a
c
t
/
E
e
q
Arithmetic average of soil moisture by Theta probes(m
3
/m
3
)
(c) Understory
2003
2004
2005
Figure 2.6: Dependence of E
act
/E
eq
on soil moisture measured by the Theta probes
28
0
0.2
0.4
0.6
0.8
1.0
E
a
c
t
/
E
e
q
(a) Overstory
0
0.5
1.0
1.5
2.0
E
a
c
t
/
E
e
q
(b) Trees
0.05 0.10 0.15 0.20 0.25 0.30 0.35 0.40
0
0.5
1.0
1.5
E
a
c
t
/
E
e
q
Arithmetic average of soil moisture by TDR (m
3
/m
3
)
(c) Understory
2003
2004
2005
Figure 2.7: Dependence of E
act
/E
eq
on interpolated soil moisture measured by the TDR
probes
29
summer drought in 2005 as demonstrated in Figure 2.2. The frequent spring rainfall events
allow the percolation of water to deeper soils that is not accessible to grasses. The open
space soil moisture at depths between 30 and 60cm in the summer of 2005 is found to be
signicantly higher compared to the summers of 2003 and 2004, while the soil moisture at
the top 30cm shows more consistent pattern of behavior over time. By taking an arithmetic
average of soil moisture at four depths up to 60cm, the soil water available to the grasses
was overestimated. In other words, an effective representation of soil water available to
the plants should account for the vertical variation in water uptake capacity limited by the
rooting depth.
An alternative to arithmetic averaging of vertical soil moisture prole is to weigh the
soil moisture at various depths by the fractions of roots present in the corresponding depths
[Baldocchi et al., 2004], i.e.,
) =
z
0
(z)dP(z)
z
0
dP(z)
, (2.5)
where P(z) is the cumulative distribution of root biomass from the surface to depth z (in
centimeters) and an exponential model based on extensive eld surveys [Jackson et al.,
1996], P(z)=1-
z
, was assumed for oak trees and grasses at the Tonzi site. The value
of the grasses was chosen to be 0.94 based on the data provided in Jackson et al. [1996],
which yields 84% of roots in the top 30cm and 13% in 30-60cm. For the oak trees, value
of 0.976 is inferred from the nding that 70% of excavated root biomass of a blue oak tree
is located above 0.5 m on average [Ishikawa and Bledsoe, 2000].
30
0
0.2
0.4
0.6
0.8
1.0
E
a
c
t
/
E
e
q
(a) Overstory
0
0.5
1.0
1.5
2.0
E
a
c
t
/
E
e
q
(b) Trees
0 0.05 0.10 0.15 0.20 0.25 0.30 0.35
0
0.5
1.0
1.5
E
a
c
t
/
E
e
q
Root density weighted soil moisture (m
3
/m
3
)
(c) Understory
2003
2004
2005
Figure 2.8: Dependence of E
act
/E
eq
on root density weighted soil moisture measured by
the TDR probes
The relationship between E
act
_
E
eq
and root density weighted representative soil mois-
ture is provided in Figure 2.8, where the pattern of the understory layer shows less inter-
annual variability compared to the cases when the arithmetic averages of soil moisture at
various depths were used. Linear decay patterns are visible for the understory and overstory
layers and for the trees. The Feddes Model can thus be applied to generalize the impact of
soil water availability on ET at the Tonzi site.
The comparisons among various representations of soil moisture at the stand scale
found that the least inter-annual variability of soil moisture effects on ET was yielded when
the root density weighted average of distributed TDR measurements were taken as the rep-
31
resentative soil moisture. This result suggests that a single-point soil moisture measurement
is not sufcient to represent the soil moisture status at a stand scale because it is not able to
capture the heterogeneity of soil moisture eld. Settin et al. [2007] reports similar ndings
through numerical studies. For this reason, spatially distributed soil moisture data collec-
tion is preferred at the stand scale. Furthermore, root biomass distribution prole plays an
important role in causing the vertical variation in water uptake capacity of the plants, and
its inuence ought to be taken into account in averaging the vertical soil moisture prole.
2.4.3 Parameterization Results and Discussions
As the applicability of the Feddes Model to characterize the effects of soil moisture
on ET appears suitable for the understory and overstory layers and for the trees in the oak
savanna ecosystem, we proceed here by identifying its parameters using WINBUGS [Lunn
et al., 2000].
In applying the WINBUGS software for inference, multiple chains of the parameters
were simulated in parallel and the modied Gelman-Rubin convergence diagnostic statis-
tics [Brooks and Gelman, 1998] were used to test the effective convergence of the samples.
Each chain had a sample size of 100,000 parameter sets with the rst 50,000 realizations
discarded to obtain a stationary distribution. One sample out of every 20 samples was se-
lected to assure the independence among samples. The resulting pool of samples was con-
sidered being drawn from the joint posterior distribution of the parameters. The marginal
distribution of each parameter can be approximated from the joint posterior samples by
32
using the kernel density estimator [Venables and Ripley, 2003].
The marginal distributions of the parameters in the Feddes Model, ,
1
and
2
(see
Figure 2.1 and equations (2.1), (2.2), and (2.3), conditioned on the root density weighted
soil moisture are given in Figure 2.9 for the overstorys, understorys and trees ET in
years 2003, 2004 and 2005. Compared to the uniform prior distributions assigned to the
parameters as shown in the Appendix, the posterior distributions are bounded by narrower
ranges with well-dened peak densities, which implies a reduction in uncertainty. Some of
the parameters, for instance,
2
for the understory layer in 2005, have multi-modal posterior
distributions, which is an advantage of the MCMC method over most traditional parameter
estimation methods.
In Figure 2.9, the inter-annual variability of the inferred parameters is evident from
the differences in their marginal distributions from year to year. In addition to the natural
variability, the inter-annual variations of model parameters could arise from the variations
in seasonal precipitation distribution patterns, because the timing and amount of a rainfall
event strongly affect how it will be partitioned, and different functional types use the avail-
able water in different ways depending on how stressed they are prior to the rainfall event
[Burgess, 2006; Rodriguez-Iturbe, 2000]. The precipitation time series given in Figure 2.2
demonstrated that 2005 has much longer raining season compared to 2003 and 2004, and
the precipitation amounts prior to the summer droughts in 2003 through 2005 are 395mm,
409mm and 717mm, respectively. It is noted that the understory layer experiences larger
variability than the trees, possibly because the grasses are less able to tackle the change in
33
0.04 0.05 0.06 0.07 0.08
0
50
100
150
200
250
300
350
P
r
o
b
a
b
i
l
i
t
y
d
e
n
s
i
t
y
0.1 0.12 0.14 0.16 0.18 0.20
0
20
40
60
80
100
120
140
(a)Overstory
0.5 0.6 0.7 0.8 0.9
0
5
10
15
20
25
30
35
0.06 0.07 0.08 0.09 0.10 0.11
0
50
100
150
200
250
300
P
r
o
b
a
b
i
l
i
t
y
d
e
n
s
i
t
y
0.1 0.12 0.14 0.16 0.18 0.20
0
20
40
60
80
100
(b)Understory
0.8 1.0 1.2 1.4
0
5
10
15
20
0.03 0.04 0.05 0.06 0.07 0.08
0
50
100
150
200
250
P
r
o
b
a
b
i
l
i
t
y
d
e
n
s
i
t
y
0.08 0.10 0.12 0.14 0.16 0.18
0
20
40
60
80
100
120
140
(c)Tree
0.6 0.8 1.0 1.2
0
2
4
6
8
10
12
14
2003
2004
2005
Figure 2.9: Posterior marginal distributions of the Feddes Model parameters
34
the ambient environment, such as change in the frequency and amount of precipitation, due
to their shallow rooting depth.
It is interesting to note that from 2003 to 2005,
1
and
2
of the understory layer were
moving closer to each other, whereas those of trees were moving in the opposite direction.
This might indicate that grasses and trees respond differently to changes in water availabil-
ity. Grasses tend to rapidly use up the available water resources, reected by lower
2
and
higher
1
, during wetter years. However, trees adopt a more conservative strategy. They
started to control ET under moister soil during wetter years so that their ET can persist into
drier soil conditions. This nding is in agreement with Rodriguez-Iturbe et al. [2001a].
The rapid depletion of water for the grasses is partly due to the fact that the supercial soil
water in the grassland evaporates and grasses have very little control over it. However, the
trees have more control over transpiration through their physiological structure. For ex-
ample, their xylem can impose hydraulic conductance limitations during period of higher
water availability to avoid dysfunction during drought [Eamus et al., 2000], which is also
known as safety-efciency trade-off [Tyree et al., 1994]. Trees can also control water loss
through the opening and closing of their stomata, which is observed on the oak trees at our
site [Xu and Baldocchi, 2003]. It is also possible that the variation in seasonal precipitation
alters the ne root distribution of the grasses and trees [Wan et al., 2002], and the effect
could be very different on trees and grasses as they compete for the same water resources.
The weighting factors used to aggregate point soil moisture to stand-level average might be
adjusted to cope with the inter-annual variability of the Feddes Model parameters. Further
35
experimental evidence is needed to address the effects of precipitation distribution pattern
on the plant responses to seasonal droughts in oak savanna.
2.4.4 Predicting ET with Calibrated Model
Each set of the sampled parameters from the MCMC method represents a realization
of the Feddes Model tted to the data. The inclusion of all realizations produced by the
algorithm denes the variability in the parameterized model, as demonstrated in Figure
2.10. Rather than identifying an optimal t of the model, the Bayesian inversion technique
with the MCMC sampling scheme yields a range of possible models, which can then be
used for statistical interpretation of the observations.
For instance, the realizations of the tted model as shown in Figure 2.10 can be used
to predict the actual ET with known meteorological variables and soil moisture. For given
conditions, each realization of the Feddes Model leads to a realization of predicted ET, and
consequently, a complete distribution of the predictions can be determined using a large
number of realizations. As an example, the model calibrated using data in 2003 is used
to predict the overstorys, understorys and trees ET between the last spring/early summer
rain and the rst winter rain in 2004 and 2005. The results are presented in Figure 2.11. The
comparisons between the predictions and the observed ET by the eddy covariance towers
reveal that the model predicts the actual ET during the dry seasons fairly well, with the
mean values being around the one-to-one line. Part of the mist is due to the inter-annual
variability of the parameters.
36
0 0.05 0.10 0.15 0.20 0.25 0.30
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
Root density weighted soil moisture (m
3
/m
3
)
E
a
c
t
/
E
e
q
Figure 2.10: Realizations of the Feddes Model compared to the data for the overstorys ET
in 2003 (Scatters are the data and lines are the realizations of the Feddes Model)
0.1 0 0.1 0.2 0.3 0.4 0.5
0.1
0
0.1
0.2
0.3
0.4
0.5
Observed ET (cm/day)
P
r
e
d
i
c
t
e
d
E
T
(
c
m
/
d
a
y
)
(a) Overstory
0.05 0 0.05 0.1 0.15 0.2
0.05
0
0.05
0.1
0.15
0.2
Observed ET (cm/day)
(b) Understory
0.1 0 0.1 0.2 0.3 0.4 0.5
0.1
0
0.1
0.2
0.3
0.4
0.5
Observed ET (cm/day)
(c) Tree
2004
2005
2004
2005
2004
2005
Figure 2.11: Prediction of ET in 2004 and 2005 with model calibrated using data in 2003
(Scatters are the mean values of the predictions and the dashed lines delineate the range
within two standard deviations from the means)
37
2.5 Conclusions
We discussed in this chapter how soil moisture controls actual evapotranspiration in a
water-limited oak savanna ecosystem located in California, and examined the applicability
of the Feddes Model for describing the effects of water stress on ET of the heterogeneous
environment as well as its components, using multi-year eld observations at the daily,
stand scale.
The inuence of soil moisture on actual ET was investigated through the dependence
of the ratio between ET, as measured by the eddy covariance towers, and the potential
ET, as approximated by the Priestley-Taylor equation, on representative soil moisture. The
observed relationship was compared to the pattern suggested by the Feddes Model in or-
der to evaluate its applicability in the oak savanna ecosystem. Various representations of
soil moisture at the stand scale, obtained from point measurements at various locations and
depths were investigated. The model parameters were inferred using a Bayesian framework
with the MCMC sampling method and their inter-annual variability was studied based on
the difference in posterior marginal distributions from year to year. Finally, the parameter-
ized model was used for predicting actual ET with uncertainty estimates determined using
the joint distribution of the parameters derived from the Bayesian framework.
The Feddes Model was found to be in agreement with the observed patterns of soil
moisture effects on ET regardless of whether the heterogeneous environment of trees and
grasses was homogenized or treated as the sum of its individual components. However,
the parameters of the Feddes Model varied with time. Grasses have lower
2
and higher
38
1
during wetter years, whereas trees have the opposite trend, i.e., they start to control ET
under moister soil (higher
2
) during wetter years so that their ET persists into drier soil
conditions (i.e., lower
1
). This inter-annual variability is primarily driven by variations in
the seasonal precipitation distribution pattern from year to year, as well as the responses of
various plant functional types to changes in soil water availability. Grasses tend to rapidly
use up the available water resources during the wetter years because they have little control
over the evaporation, whereas trees are able to control water loss by limiting the xylem
hydraulic conductance and by controlling their stomata. This difference in responses can
also be a result of change in ne root distributions due to water availability in early seasons.
The selection of representative soil moisture leads to considerable difference in the
inter-annual variability of model parameters. Among the various averaging schemes tried
in this study, including (1) the arithmetic average of soil moisture measurements at different
depths obtained at one location, (2) the arithmetic average of the spatially distributed soil
moisture measurements at different depths and (3) the root density weighted average of the
distributed soil moisture measurements at different depths, the last scheme yielded the most
consistent behavior of the Feddes Model over time. The study demonstrates that distributed
sampling of soil moisture is necessary to study the effects of soil moisture availability
on ET in open canopy ecosystems. Furthermore, the soil water availability at different
depths has a different inuence on the total ET depending on the fraction of root biomass
present at the corresponding depths. This difference can be accounted for by using the root
density weighted average of the vertical soil moisture prole to produce representative soil
39
moisture.
The inter-annual variability of climate brings inherent variability to model parameters
in addition to parameter uncertainties arising from measurement errors and an incomplete
understanding of the underlying physical mechanisms. It is thus insufcient to assign deter-
ministic values to model parameters without identifying the associated uncertainties. The
parameter estimation methodology presented in this study, a Bayesian framework based
on the MCMC sampling method, provides a systematic tool to quantify the parameter un-
certainties conditioned on eld observations and to directly estimate the prediction uncer-
tainties using the model parameterized in this way. We calibrated the Feddes Model using
data from 2003 and predicted the actual ET in 2004 and 2005 with quantied uncertainty.
The comparison between the predicted and measured ET showed good agreement. The
calibrated Feddes Model can be incorporated into the water balance equation to study wa-
ter dynamics at the stand scale and eventually to better understand the water cycle in a
heterogeneous environment.
This study developed a methodology to model the changes in ecosystem ET under
drought conditions and to statistically parameterize the model using eld data. The suc-
cessful application of the methodology in the complex oak savanna ecosystem inspires its
employment in other water-limited ecosystems.
40
Chapter 3
Theoretical Analysis for Sap Flow
Measurements using Heat Ratio Method
3.1 Introduction
Heat pulse method is widely employed to measure water use by trees [Cohen et al.,
1981; Granier, 1985; Swanson, 1994; Smith and Allen, 1996; Green et al., 2003]. It works
by deducing sap ow rate from the speed at which a short heat pulse is propagated through
the porous medium. The heat pulse technique has the advantages of simple instrumentation
and lower power consumption. Among the numerous variations of the heat pulse technique,
heat ratio method (HRM) introduced by Burgess et al. [1998] has been widely applied in
various elds due to its ability to detect reverse and low ow rates and its straightforward
instrumentation [Burgess et al., 2001]. The apparatus for the HRM consists of a pair of
41
upstream
thermocouple
heater
downstream
thermocouple
d
x
u
x
heartwood sapwood
bark
V
x
z
Figure 3.1: Diagram of heat ratio apparatus
thermocouple probes located equidistantly upstream and downstream from a heating ele-
ment as depicted in Figure 3.1. The probes are parallel and aligned in a common plane.
The heat pulse velocity in HRM is deduced from the ratio of temperature increases
recorded at downstream and upstream temperature probes, following the release of a heat
pulse by the central heating element. The theoretical basis underlying HRM is an analyti-
cal solution to the heat transport equation developed by Marshall [1958], which is derived
under idealized assumptions. For instance, the porous medium is assumed unbounded and
homogeneous, and heat is instantaneously released from an innite line source. However,
these assumptions are routinely violated in real applications of HRM, e.g., tree sapwood
and heating element are of nite dimension rather than being innite, and heat is not re-
42
leased instantaneously but over a duration of time. The departure of the real conditions
from the ideal assumptions may result in different temperature eld around the heating
element, and consequently affect the theoretical basis of HRM.
The objectives of this study are to (1) develop a solution to the heat transport process
in tree sapwood with more realistic assumptions, (2) compare the difference in temperature
elds approximated by Marshalls idealized solution and our more realistic solution, and (3)
provide modications if necessary to the fundamental equation of HRM based on improved
understanding of the heat transport process.
3.2 Theory
3.2.1 Heat transport equation in sapwood
Wood is considered as a porous medium composed of cellulose, a solid fraction forming
the vessel walls, and sap, a liquid lling the void space. The heat released from the central
heating element is transported away by conduction and convection. Assuming sap water is
moving uniformly in the xdirection and wood is thermally homogeneous and isotropic,
the heat transfer process can be described as [Carslaw and Jaeger, 1959; Gribben, 1999]
T
t
=
_
2
T
x
2
+
2
T
y
2
+
2
T
z
2
_
v
h
T
x
+Q(x)(y)(z)(t) (3.1)
where T is temperature rise, t is time, is the thermal diffusivity of the wood matrix, and
x, y, and z are space coordinates, Q represents the heat source term, and () is the Dirac
43
function. The heat pulse velocity, v
h
, is related to sap velocity, v
s
, by the following equation
dened by Marshall [1958],
av
s
=
w
c
w
s
c
s
v
h
(3.2)
where a is the fraction of the cross-sectional area of conducting sapwood occupied by
moving sap streams and and c are density and specic heat capacity, with the subscript s
referring to sap and w referring to wood matrix. The volumetric sap ux density per unit
cross-sectional area of sapwood is given by J
s
= av
s
.
3.2.2 Marshalls solution
Marshall [1958] derives an analytical solution to Eq. (3.1) considering instantaneous
heat released from an innite line source normal to the x y plane and passes through the
point (x, y) = (0, 0), in an innite medium at uniform initial temperature, which is
T(x, y, t) =
q
4ct
exp
_
(x v
h
t)
2
+y
2
4t
_
(3.3)
where q is the quantity of heat released per unit length.
If temperature increase is measured at points equidistant downstream and upstream
from the line source, the heat pulse velocity can be derived from Eq. (3.3) as
v
h
=
x
ln
T
d
T
u
(3.4)
where x is the distance between the heater and either temperature probe, T
d
and T
u
are
temperature increases measured at x cm downstream and upstream from the heater, respec-
44
0 20 40 60 80 100 120
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
Time after release of heat pulse (s)
T
e
m
p
e
r
a
t
u
r
e
c
h
a
n
g
e
(
o
C
)
0 20 40 60 80 100 120
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
2.2
2.4
Time after release of heat pulse (s)
l
n
(
T
d
/
T
u
)
Figure 3.2: Example of heat response curves taken downstream and upstream from the
heater probe and the corresponding time series of log-ratio of temperature increases. The
dashed line in the right panel is the log-ratio calculated from average temperature rise over
t=60-100s following the heat pulse.
tively. Eq. (3.4) is the theoretical basis of HRM [Burgess et al., 2001]. An example of
temperature increases measured using the HRM apparatus and the calculated temperature
increase ratios is shown in Figure 3.2.
3.2.3 Improved solutions
Ren et al. [2000] present another solution for Eq. (3.1) based on the same assumptions
as Marshalls except that the instantaneous heating is replaced by a heat pulse lasting from
0 to t
0
, which is a better representation of the heat source in practice. Their solution is
45
T(x, y, t) =
q
/
4c
t
tt
0
s
1
exp
_
(x v
h
s)
2
+y
2
4s
_
ds (3.5)
where q
/
is the quantity of heat released per unit length per unit time and t
0
is the duration
of heat pulse. This solution is for t >t
0
.
It is further studied in Wang et al. [2002] that Eq.(3.5) can also lead to eq. (3.4) when the
temperature probes are aligned symmetrically about the heater probe. If asymmetry occurs,
Eq. (3.4) asymptotically approaches the heat pulse velocity using average distance between
the probes as x. Therefore, it is recommended that the ratios of temperature increases
should be calculated at a time when its value has reached a constant.
However, the assumptions of an innite medium and an innite line source correspond-
ing to Marshlls and Rens solutions are not satised in sap ow applications. To account
for the nite width of the sapwood, Gribben [1999] developed a solution for the tempera-
ture increases following an instantaneous heat release from a linear line source as
T(x, y, z, t) =
q
ct
exp
_
(x v
h
s)
2
+y
2
4t
_
n=1
sin(b
n
L)cos(b
n
z)
2b
n
L+sin(2b
n
L)
exp
_
b
n
2
t
_
(3.6)
where L is the depth of the sapwood and the line source is assumed to extend through the
entire sapwood depth, and b
n
is obtained by solving the equation, tan(bL) = K/b, with
(n1)/L < b
n
< (2n1)/(2L).
In deriving Eq. (3.6), it is assumed that geometry in the vicinity of the heating element
is at since length of heating element is much smaller than the tree radius, and no heat ow
boundary and Newtons Law of Cooling are applied at the bark (z = 0) and at the boundary
46
between sapwood and heartwood (z = L), respectively, i.e.,
T
z
= 0 at z = 0 (3.7)
T
z
+KT = 0 at z = L (3.8)
where K is the parameter describes how fast the surface cools and it is not expected to be
known because the solution is not sensitive to its values [Gribben, 1999].
Based on Gribbens solution, we further replace the instantaneous heat source with
a pulsed one, and add the exibility that the length of line source can be shorter than
the sapwood depth. Skipping the evolving steps, our nal solution based on the realistic
assumptions is obtained as follows,
T(x, y, z, t) =
q
/
c
n=1
sin(b
n
a)cos(b
n
z)
2b
n
L+sin(2b
n
L)
t
tt
0
s
1
exp
_
(x v
h
s)
2
+y
2
4s
b
n
2
s
_
ds
(3.9)
where a is the length of line source. To ensure the no heat ow boundary at the tree bark,
it is recommended to insulate the tree bark around the sap ow apparatus in eld.
In all the solutions presented here, sap is assumed to ow uniformly through the porous
medium that is at uniform initial temperature. In addition, the ow deformation caused by
the implanted probes is not considered, assuming that the size of the probes is innitesi-
mally small compared to the tree size.
47
0 20 40 60 80 100 120
0.5
1
1.5
2
2.5
3
3.5
Time after release of heat pulse (s)
T
e
m
p
e
r
a
t
u
r
e
c
h
a
n
g
e
(
o
C
)
=0.003cm
2
/s, v
h
=30.0cm/hr, x=0.6cm
0 20 40 60 80 100 120
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
Time after release of heat pulse (s)
T
e
m
p
e
r
a
t
u
r
e
c
h
a
n
g
e
(
o
C
)
=0.003cm
2
/s, v
h
=30.0cm/hr, x=0.6cm
Marshall
Ren
Chen
Marshall
Ren
Chen
Figure 3.3: The temperature increases calculated from various theoretical models. Our
solution is labeled as Chens
3.3 Results and Discussions
3.3.1 Comparison of calculated temperature elds
It is demonstrated in the previous section that the various solutions to heat transport pro-
cess differ in their fundamental assumptions, which may consequently lead to differences
in their calculated temperature elds around the line source. Figure 3.3 shows an example
of temperature rises calculated from various theoretical models at 0.6 cm downstream (x =
0.6 cm) and upstream (x = -0.6 cm) from the line source assuming = 0.0030 cm
2
/s, v
h
=
30 cm/hr, y = 0.0 cm, z = 1.0 cm, L = 7.0 cm, a = 3.0 cm, and t
0
= 6 s. It is evident from
this example that signicant differences among the solutions mostly occur at early times,
say, before the arrival of peak, while the differences at late times are negligible.
However, the differences among these solutions may vary with probe geometry, wood
thermal diffusivity, heat pulse velocity as well as the sapwood depth. Therefore, an in-depth
48
investigation is conducted on how the aforementioned parameters affect the difference in
calculated temperature rises caused by the theoretical models. As the largest difference is
anticipated between the Marshalls solution and our solution, the comparisons are focused
on these two models, and their relative difference is calculated as [
_
T
Marshall
T
Chen
_
/T
Chen
[.
Inuence of vertical probe distance (x)) and heat pulse velocity (v
h
)
Assuming values of , y and z are xed at 0.0030 cm
2
/s, 0.0 cmand 1.0 cm, respectively,
the change of relative difference in temperature rise in sapwood with a depth of 7 cm is
shown in Figure 3.4 as a function of time and x-locations. The difference is found to
be symmetric about the x values. Therefore, only results for positive x values are provided
here. It is observed that most of the signicant differences occur in the early times for all the
cases represented in the plots. In addition, the early-time difference is observed to increase
with probe spacing in x-direction. Nevertheless, this trend can be reversed around the time
approaching the minimum difference, which is dependent on both the probe spacing and
heat pulse velocity.
It is evident in Figure 3.4 that, at a xed location, the heat pulse velocity affects the de-
creasing rate of difference in the early times, the time for the difference to reach its asymp-
totic value, as well as the magnitude of the asymptotic difference. The example shown in
Figure 3.5, extracted from Figure 3.4 for x = 0.6 cm, exhibits that the difference decreases
with heat pulse velocity in early times, whereas the asymptotic difference increases with
the heat pulse velocity. It is noticed that, for t >40 s, the difference corresponding to any
49
0 50 100 150 200
10
4
10
2
10
0
10
2
Time(s)
D
i
f
f
e
r
e
n
c
e
i
n
t
e
m
p
e
r
a
t
u
r
e
r
i
s
e
v
h
= 0.0 cm/hr
x=0.4
x=0.6
x=0.8
x=1
x=1.5
0 50 100 150 200
10
4
10
2
10
0
10
2
Time(s)
D
i
f
f
e
r
e
n
c
e
i
n
t
e
m
p
e
r
a
t
u
r
e
r
i
s
e
v
h
=10.0 cm/hr
0 50 100 150 200
10
4
10
2
10
0
10
2
Time(s)
D
i
f
f
e
r
e
n
c
e
i
n
t
e
m
p
e
r
a
t
u
r
e
r
i
s
e
v
h
=30.0 cm/hr
0 50 100 150 200
10
4
10
2
10
0
10
2
Time(s)
D
i
f
f
e
r
e
n
c
e
i
n
t
e
m
p
e
r
a
t
u
r
e
r
i
s
e
v
h
=50.0 cm/hr
Figure 3.4: Difference in temperature rise between Marshalls solution and our solution as
affected by xlocation and heat pulse velocity for L = 7 cm, = 0.0030 cm
2
/s, y = 0 cm,
and z = 1.0 cm. The difference is calculated as [
_
T
Marshall
T
Chen
_
/T
Chen
[.
50
0 20 40 60 80 100 120 140 160 180 200
10
4
10
3
10
2
10
1
10
0
10
1
Time(s)
D
i
f
f
e
r
e
n
c
e
i
n
t
e
m
p
e
r
a
t
u
r
e
r
i
s
e
v
h
=0.0
v
h
=10.0
v
h
=30.0
v
h
=50.0
Figure 3.5: Difference in temperature rise between Marshalls solution and our solution as
affected by heat pulse velocity for L = 7 cm, = 0.0030 cm
2
/s, x = 0.6 cm, z = 1.0 cm, and
v
h
= 30 cm/hr. The difference is calculated as [
_
T
Marshall
T
Chen
_
/T
Chen
[.
heat pulse velocity is less than 3%, which may be considered negligible.
In addition, it is noted that the minimum differences in the plots are not the exact ones,
which should be zero ( in log-scale) when two solutions meet. Nevertheless, the time
corresponding to the minimum difference is close to the true ones. Except for these cases
corresponding to small values of x and v
h
, the solutions of Marshalls model and our model
meet only once after the arrival of peak.
51
0 20 40 60 80 100 120 140 160 180 200
10
5
10
4
10
3
10
2
10
1
10
0
10
1
Time(s)
D
i
f
f
e
r
e
n
c
e
i
n
t
e
m
p
e
r
a
t
u
r
e
r
i
s
e
y=0.0
y=0.2
y=0.4
y=0.6
y=0.8
y=1.0
Figure 3.6: Difference in temperature rise between Marshalls solution and our solution as
affected by y-coordinate for L = 7 cm, = 0.0030 cm
2
/s, x = 0.6 cm, z = 1.0 cm, and v
h
=
30 cm/hr.The difference is calculated as [
_
T
Marshall
T
Chen
_
/T
Chen
[.
Inuence of lateral position (y)
In both Marshalls solution and our solution, y-coordinate of the measurement point
is a necessary component to derive its temperature rise. As revealed in Figure 3.6, the
difference of temperature rise between two solutions is inuenced by values of y. With the
other parameters being xed, the difference at early times increases with the y value, while
same asymptotic value will be reached when elapsed time is sufciently long. At t >60 s,
the difference is within 2% and 10%, respectively, for y 0.6 cm or otherwise.
52
0 20 40 60 80 100 120 140 160 180 200
10
5
10
4
10
3
10
2
10
1
10
0
10
1
Time(s)
D
i
f
f
e
r
e
n
c
e
i
n
t
e
m
p
e
r
a
t
u
r
e
r
i
s
e
z=1.0
z=1.5
z=2.5
Figure 3.7: Difference in temperature rise between Marshalls solution and our solution as
affected by y-coordinate for L = 7 cm, = 0.0030 cm
2
/s, x = 0.6 cm, y = 0.0 cm, and v
h
=
30 cm/hr.The difference is calculated as [
_
T
Marshall
T
Chen
_
/T
Chen
[.
Inuence of radial depth (z)
One of the key differences between two solutions is that Marshalls solution is two-
dimensional, while ours is three-dimensional. The coordinate along z-direction represents
the relative position to nite line source and the distance to boundaries. Therefore, the
impact of z on the difference of two solutions may be more complex than x and y due to
the interacting effects of boundary conditions and nite length of line source. The example
provided in Figure 3.7 proves that there is no clear trend of the difference against z. The
largest difference is yielded at the mid-point of the line source at z = 1.5 cm when t > 40 s,
whereas the difference in the same time frame for other z values is smaller.
53
0 20 40 60 80 100 120 140 160 180 200
10
4
10
3
10
2
10
1
10
0
10
1
Time(s)
D
i
f
f
e
r
e
n
c
e
i
n
t
e
m
p
e
r
a
t
u
r
e
r
i
s
e
L=5.0
L=7.0
L=9.0
L=11.0
Figure 3.8: Difference in temperature rise between Marshalls solution and our solution as
affected by the sapwood depth for = 0.0030 cm
2
/s, x = 0.6 cm, y = 0.0 cm, z = 1.0 cm,
and v
h
= 30 cm/hr. The difference is calculated as [
_
T
Marshall
T
Chen
_
/T
Chen
[.
Inuence of sapwood depth (L)
The inuence of sapwood depth on the solution difference is related to the assumptions
of innite medium and innite line source in Marshalls solution. For a typical range of
sapwood depth from 5 cm to 11 cm, the example presented in Figure 3.8 illustrates that
the asymptotic difference at t > 60 s increases with sapwood depth, while the decay of
difference with time is very slow for small sapwood depth, e.g., L = 5.0 cm, compared to
the other depths considered. The reason may be that boundary effects is more dominant in
small sapwood depth, whereas the inappropriate assumption of innite line source plays a
more signicant role in medium to large sapwood depths.
54
0 20 40 60 80 100 120 140 160 180 200
10
4
10
3
10
2
10
1
10
0
10
1
Time(s)
D
i
f
f
e
r
e
n
c
e
i
n
t
e
m
p
e
r
a
t
u
r
e
r
i
s
e
=0.0025 cm
2
/s
=0.0030 cm
2
/s
=0.0035 cm
2
/s
=0.0040 cm
2
/s
Figure 3.9: Difference in temperature rise between Marshalls solution and our solution as
affected by the sapwood depth for L = 7 cm, x = 0.6 cm, y = 0.0 cm, z = 1.0 cm, and v
h
=
30 cm/hr. The difference is calculated as [
_
T
Marshall
T
Chen
_
/T
Chen
[.
Inuence of thermal diffusivity ()
An example showing the impact of on solution difference is given in Figure 3.9. The
range of value is chosen based on the calculation provided in Appendix D, which shows
that the wood thermal diffusivity at the site may vary between 0.0025 and 0.004 cm
2
/s in
response to various wood densities and moisture conditions. Larger difference is observed
in Figure 3.9 for smaller value in early times. However, the trend is reversed for t > 50
s, and at the same time, the difference caused by varying value is diminishing with time.
55
3.3.2 Impacts on HRM
It is of interest to investigate whether the estimation of the heat pulse velocity based
on our improved solution differs from that based on Marshalls solution. As presented in
the previous section, the difference between two solutions is normally negligible at late
times. It can be expected that Eq. (3.4) is still valid for symmetric probe alignment if
the temperature increase are recorded at late times, say over 60-100s following the heat
pulse. It is conrmed true using the extensive data for comparing solution difference in the
preceding section.
Nevertheless,the probe geometry in practice can depart from the ideal alignment as-
sumed to derive Eq. (3.4): the probe spacing may not be symmetric in x-direction and the
three probes outlined in Figure 3.1 may not lie in the same plane in y-direction. In order
to examine the applicability of Eq. (3.4) under arbitrary probe alignments, the data in the
previous section are used to create numerous combinations of probe geometry and the heat
pulse velocity is found to be linearly related to
x
ln
T
d
T
u
, i.e.,
v
h
= B
0
+B
1
x
ln
T
d
T
u
(3.10)
where B
0
and B
1
are empirical factors, x is the average distance of downstream and up-
stream temperature probes from the line source, and T
d
and T
u
are average temperature
increases over 60s t 100s measured by the downstream and upstream temperature
probes, respectively. The average temperature increase is used to calculate the ratio in the
consideration that temperature increase is usually small for t > 60s, especially at the up-
56
stream probe. As a result, random noise in individual temperature measurement may lead
to unreliable result of ratio.
Further study shows that B
0
and B
1
are mainly controlled by the geometry asymmetry,
while the other parameters, including , L, and z, have no signicant impact. The contour
plots in Figure 3.10 demonstrate that B
0
and B
1
are nearly linearly related to x
d
x
u
and
y
2
u
y
2
d
, while there is no noticeable difference caused by varying z values. It is also noted
from the plots that B
1
only varies slightly around 1, whereas B
0
can be as large as 30 cm/hr.
Linear regression of B
0
and B
1
on probe geometry along with , L, and z conrms that
, L, and z are not signicant factors, and the regression equations are obtained as
B
0
= 0.031+22.35x
d
21.91x
u
44.77
_
y
2
u
y
2
d
_
0.42x
d
x
u
+26.55x
d
_
y
2
u
y
2
d
_
+27.39x
u
_
y
2
u
y
2
d
_
19.77x
d
x
u
_
y
2
u
y
2
d
_
(3.11)
B
1
=
_
100+1.32x
d
1.30x
u
2.66
_
y
2
u
y
2
d
_
2.10x
d
x
u
+1.58x
d
_
y
2
u
y
2
d
_
+1.63x
u
_
y
2
u
y
2
d
_
1.18x
d
x
u
_
y
2
u
y
2
d
_
10
2
(3.12)
where x
d
and x
u
are both taken as their absolute values. Both equations yield R
2
value of
0.999, which is a sign of good tting to the data.
3.4 Conclusions
This study re-evaluated the theoretical basis of sap ow measurements using HRM.
The fundamental equation of HRM was originally derived from Marshalls solution to heat
57
x
d
x
u
y
u
2
y
d
2
B
0
(
c
m
/
h
r
)
w
i
t
h
L
=
7
c
m
,
z
=
1
c
m
0
.
6
0
.
4
0
.
2
0
0
.
2
0
.
4
0
.
6
0
.
5 0
0
.
5 1
x
d
x
u
y
u
2
y
d
2
B
1
w
i
t
h
L
=
7
c
m
,
z
=
1
c
m
0
.
6
0
.
4
0
.
2
0
0
.
2
0
.
4
0
.
6
0
.
5 0
0
.
5 1
x
d
x
u
y
u
2
y
d
2
B
0
(
c
m
/
h
r
)
w
i
t
h
L
=
7
c
m
,
z
=
2
.
5
c
m
0
.
6
0
.
4
0
.
2
0
0
.
2
0
.
4
0
.
6
0
.
5 0
0
.
5 1
x
d
x
u
y
u
2
y
d
2
B
1
w
i
t
h
L
=
7
c
m
,
z
=
2
.
5
c
m
0
.
6
0
.
4
0
.
2
0
0
.
2
0
.
4
0
.
6
0
.
5 0
0
.
5 1
0
.
9
8
5
0
.
9
9
0
.
9
9
5
1 1
.
0
0
5
1
.
0
1
2
0
1
0
0 1
0
2
0
0
.
9
8
5
0
.
9
9
0
.
9
9
5
1 1
.
0
0
5
1
.
0
1
2
0
1
0
0 1
0
2
0
Figure 3.10: Contours of empirical factors versus spacing asymmetry in x- and y-
directions.
58
transport process in sapwood with idealized assumptions. we developed an improved solu-
tion to the same process with more realistic assumptions. More specically, the sapwood
was considered bounded porous medium, the line source had nite length, which may or
may not extend through the entire sapwood depth, and the heat was released from the line
source over a duration. Extensive comparisons on the difference of calculated temperature
elds by Marshalls solution and our solution revealed that most signicant discrepancy
occurs around the early times, whereas the difference is negligible at later times, say t >
60 s. It is therefore recommended that when employing HRM the temperature increases
should be measured in this late time window.
Furthermore, the fundamental equation of HRM was derived on the basis of symmetric
probe alignment. In practice, however, the probes can be misaligned during the installation
process, even if the probes were carefully installed with a drilling guide. Due to this prac-
tical consideration, this study also investigated whether the fundamental equation of HRM
was changed under asymmetric probe alignments. With the numerical data generated from
our solution, which cover a wide range of possible sapwood properties and probe align-
ments, a signicant shift from the true heat pulse velocity may be resulted from the asym-
metric alignment if the heat pulse velocity is estimated by the HRM equation for symmetric
probe alignment. Therefore, the fundamental equation of HRM can be slightly changed to
include an interception (B
0
) and slope (B
1
) to account for this asymmetry. Linear regres-
sion of B
0
and B
1
on sapwood properties and probe alignment found that B
0
and B
1
can be
accurately determined by probe geometry in x- and y- directions. The implementation of
59
this revised HRM equation is shown in next chapter.
This study not only improves our understanding of the underlying principles of HRM,
but also provides sounding evidence that the fundamental equation of HRM can only be
slightly adjusted to account for the departure of real conditions from the idealized one.
These ndings are the solid base for our choice of HRM in monitoring tree transpiration.
60
Chapter 4
On Determining Wood Thermal
Diffusivity and Probe Geometry Using
In-Situ Heat Response Curves for Sap
Flow Measurements
4.1 Introduction
The sap ow method has long been used to measure transpiration by individual plants
[Barrett et al., 1995; Burgess et al., 2001; Cermak et al., 1973; Chandra et al., 1994;
Kluitenberg and Ham, 2004; Marshall, 1958; Swanson and Lee, 1966], and has a history
of applications in hydrology, forestry, ecology, agriculture, and horticulture [Cohen et al.,
61
1988; Cohen and Li, 1996; Eastham and Gray, 1998; Fernandez et al., 2001; Wilson et al.,
2001]. The transpiration measurement at individual tree by the sap ow technique not only
provides sounding evidence to study the plant response to ambient stresses or disturbances
such as drought and irrigation [Cermak et al., 1993; Duursma et al., 2008; Dye, 1996;
Oren et al., 1999; Poyatos et al., 2008; West et al., 2008], it can also be upscaled to areal
transpiration [Cermak et al., 2004; Cienciala et al., 1999; Crosbie et al., 2007; Granier
et al., 1996; Saugier et al., 1997; Whitley et al., 2008]. The sap ow method has contributed
signicantly to studies of nighttime transpiration [Dawson et al., 2007; Fisher et al., 2007]
and hydraulic redistribution in root zone [Burgess et al., 2000; Kurz-Besson et al., 2006;
Nadezhdina et al., 2006; Scott et al., 2008], both of which are important to understanding
plant water use in semiarid regions. A recent special issue in Plant and Soil [Burgess,
2008] provides a comprehensive review of new developments and applications of the sap
ow method.
In general, there are three major thermometric techniques used to measure sap ow:
heat pulse, heat balance, and heat dissipation. In all the three techniques heat is used as a
tracer but in different ways: With the heat pulse technique, sap ow rate is measured by
determining the speed at which a short heat pulse is propagated through the porous medium;
With the heat balance technique, sap ow rate is determined from the vertical heat loss by
convection, which is calculated from the balance of heat uxes into and out of the heated
stem section. Heat can either be externally applied to the entire circumference of the trunk
or be internally applied to a segment of the trunk on large trees. Finally, with the thermal
62
dissipation technique, which is often known as the Granier method [Granier, 1985], an
empirical equation is used to relate the sap ow rate and temperature difference between
two thermocouple probes that are about 10 cm apart, and constant power is applied to the
heater that is put together with the thermocouple probe on the top. We refer readers to the
work of Smith and Allen [1996] for a review of the underlying theories of these techniques
and to Swanson [1994] for a review of the history of their use.
This study focuses on the heat pulse technique, which has advantages of low power
consumption, simple instrumentation and automated data collection. Green et al. [2003]
provide a detailed review of its theory and practical applications. Among the variations of
the heat pulse technique, the heat ratio method introduced by Burgess et al. [1998] has been
widely applied due to its ability to detect reverse and low ow rates and its straightforward
instrumentation [Burgess et al., 2001]. The apparatus for the heat ratio method consists
of a pair of thermocouple probes located equidistantly upstream and downstream from a
heating element as shown in Figure 3.1.
When measuring the sap velocity using the heat ratio method, it is critical to know the
thermal diffusivity of the wood matrix and the distance of the thermocouple probes from
the heater probe. While thermal diffusivity may be estimated from the core samples of the
sapwood [e.g., Scott et al., 2008] or calculated from measurements of stem water content
and sapwood density [e.g., Burgess et al., 2001], many studies do not report how thermal
diffusivity values are chosen. Given the exact probe spacing, the wood thermal diffusiv-
ity can be estimated using the methodology developed by the soil science community to
63
determine soil heat properties using the heat pulse technique [Bristow et al., 2001, 1994;
Campbell et al., 1991]. Nevertheless, the exact spacing and geometry of the probes cannot
be guaranteed in practice even when the probes are carefully installed, due to the nature of
the wood matrix. This problem is especially prevalent in hardwood species such as oak and
maple. Considering the fact that the estimation of sap ow rates is sensitive to the probe
spacing, e.g., a 1-mm error in probe spacing can cause more than 10% difference in the
estimated sap ow rates, Burgess et al. [2001] suggest a correction procedure for the probe
misalignment. However, it is dependent on obtaining zero-ow conditions, which is either
assumed to happen at night or is achieved by cutting down trees. Therefore, there is a need
for in-situ and non-destructive approach to determine both the wood thermal properties and
probe geometry, in order to improve the accuracy of the sap ow measurements with the
heat ratio method.
The objective of our study is to provide a systematic, non-destructive, and replicable
methodology to determine wood thermal diffusivity and probe geometry for the sap ow
measurements using the heat ratio apparatus. The estimation of these parameters is condi-
tioned on the time series of temperature increases, i.e., temperature response curves, mon-
itored by the downstream and upstream temperature probes after a heat pulse is released
by the heater probe. The temperature response curves are controlled by the combination of
thermal diffusivity, probe geometry, and convective velocity, which is in turn a function of
wood thermal diffusivity, probe geometry, and the ratio of temperature increases detected
by the downstream and upstream probes. Thus the wood thermal diffusivity and probe
64
geometry may be inversely estimated provided the temperature response curves taken at
downstream and upstream locations from the heater probe. The primary advantage of the
methodology is that it relies on the information that can be obtained using the installed
probes without any further disturbance to the tree.
The difculties in parameter estimation procedure may arise from various sources,
among which the most important one is the uncertainties rooted in the measurement error
of the temperatures and the inadequacy of selected model to approximate the heat trans-
port process in wood stem. Therefore, a Bayesian inversion framework with Markov Chain
Monte Carlo (MCMC) sampling scheme is chosen for its strengths in dealing with uncer-
tainties arising from various sources and in combining prior knowledge of the parameters
to be estimated [Clark, 2005; Clark and Gelfand, 2006]. Empirical factors are adopted to
account for the departure of the eld conditions from the idealized conditions that form
the theoretical basis of the heat ratio method, and they are estimated together with wood
thermal diffusivity and probe geometry.
4.2 Materials and Methods
4.2.1 Data acquisition
The heat ratio apparatus employed in this study consists of three 3-cm-long hypodermic
needles, two of which are temperature probes and one is the heater probe with a resistance
around 20 ohms. For each temperature probe, there are two thermocouple junctions in-
65
stalled at two different depths, 1 cm and 2.5 cm into the sapwood, respectively, to capture
the radial variation of sap ow. The upstream and downstream temperature probes were
installed to as close to 0.6 cm equidistantly from the central heater probe as possible. Each
tree had two sets of sensors, one at breast height (around 1.5 m from the ground) and the
other close to ground (around 0.3 m from the ground). Both sets were installed on the
north side of the tree when possible to minimize the inuence of diurnal bole temperature
uctuation, and all sensors were insulated with aluminum foil.
For each test, a 12-15 V voltage was applied on the heater probe for six seconds to gen-
erate the heat pulse. Temperature traces were recorded every two seconds until 100 s after
the heat pulse stopped. The temperature increases were calculated by subtracting the initial
temperature taken before the onset of the heat pulse from the temperature traces. The test
was repeated in different seasons because wood thermal diffusivity may vary throughout
the year due to the change of stem water content and ambient temperature.
4.2.2 Parameter Estimation Method
It has been investigated in Chapter 3 that the heat pulse velocity is linearly related to the
ratio of temperature increases recorded between 60-100 s following the heat pulse, with the
empirical factors determined by the probe geometry. Asymmetry in probe geometry usually
results in a shift in the estimation of heat pulse velocity. In addition to such correction for
non-ideal probe geometry, ow deformation imposed by the implanted probes is found
to underestimate the heat pulse velocity using heat pulse technique [Burgess et al., 2001;
66
Swanson and Whiteld, 1981]. Empirical factors are thus introduced to correct for the
wounding effects as well as the difference in thermal properties of wood and probe material.
It is studied in Burgess et al. [2001] that a linear correction function performs as well as a
polynomial one. Therefore, we adopt a linear correction factor, W, then the corrected heat
pulse velocity is v
ch
=Wv
h
with v
h
determined by equation (3.10).
Reliable sap ow estimation using HRM relies on the determination of appropriate pa-
rameter values. Here we discuss how to determine the parameters to calculate v
h
: empirical
factors (B
0
and B
1
), wood matrix thermal diffusivity (), and the geometry of probe instal-
lation (x
d
, x
u
, y
d
, and y
u
), while the estimation of wounding correction factor (W) is not
considered. The parameter estimation is conditioned on the heat response curves measured
by the downstream and upstream temperature probes following the release of a heat pulse.
Given a set of heat response curves taken downstream and upstream as shown in Figure
3.2, it is assumed that they can be described by the Marshalls analytical solution with
the heat pulse velocity determined from Eq. (3.10). The ratio of temperature increases
downstream and upstream is calculated from the temperature increases measured over 60-
100s following the heat pulse, and a noise is added to simulate the measurement error in
practice. Then, the parameters are selected on the basis of model tting to the time series
data.
In order to deal with the uncertainty involved in measured heat response curves and
to facilitate the proceeding uncertainty analysis using the derived parameters, Bayesian
inversion technique with the Markov Chain Monte Carlo (MCMC) sampling method is
67
adopted for parameter estimation. This framework is implemented in the WINBUGS soft-
ware [Lunn et al., 2000], which is a powerful tool for Bayesian inverse modeling with open
access. A more detailed description of the estimation procedure can be found in Chen et al.
[2008b]. Multiple chains of the parameters were simulated in parallel and the modied
Gelman-Rubin convergence diagnostic statistics [Brooks and Gelman, 1998] were used to
test the effective convergence of the samples. Each chain had a sample size of 45,000 pa-
rameter sets with the rst 40,000 realizations discarded to obtain a stationary distribution.
One sample out of every ten was selected to assure the independence among samples. The
resulting pool of samples was considered realizations of the joint posterior distribution of
the parameters. The marginal distribution of each parameter was approximated from the
joint posterior samples by using the kernel density estimator [Venables and Ripley, 2003].
Details in MCMC implementation can be found in the appendix.
4.3 Results and Discussions
4.3.1 Model verication
The performance of the proposed parameter estimation framework depends on two im-
portant factors: one is whether Marshalls analytical solution is appropriate to describe the
heat transport process in sapwood for a specic application; the other is whether the pro-
posed framework is able to nd the true parameter values if no model error is present. The
latter can be tested using synthetic response curves generated from Eq. (3.3) with known
68
, v
h
, x
d
, x
u
, y
d
, and y
u
. The parameters estimated using our proposed framework can then
be compared to their true values. Furthermore, the sensitivity of estimated parameters to
their prior knowledge can be investigated using the synthetic data.
Marshall [1958] mentions that the solution to a given heat response curve may not be
unique, i.e., different combinations of , v
h
and x may produce the identical heat response
curves. To overcome this problem, heat response curves from multiple tests are used rather
than a single curve. Each test consists of a pair of curves, as shown in Figure 3.2, which
share same and v
h
. All the tests are conducted on the same set of probes, i.e., they have
same x
d
, x
u
, y
d
, and y
u
. The heat pulse velocity corresponding to each test is determined by
and x through Eq. (3.10). With these extra constraints, we anticipate to achieve parameter
distributions centered around their true values if no model error is present.
The synthetic case to verify the parameter estimation framework contains three tests
for a probe set with x
d
= 0.5 cm, x
u
= 0.7cm, y
d
= 0.1 cm, and y
u
= 0.2 cm. The thermal
diffusivity for each test is 0.0030, 0.0035, and 0.0028 cm
2
/s, and the heat pulse velocity for
each test is 30, 15, and 10 cm/hr, respectively. The heat released by the heater is set at 12
J/cm. The density and specic heat capacity of fresh wood are set at 1530 kg/m
3
and 1500
J/(kg
o
C), respectively. In this formulation, x
u
is negative because the x-coordinate of Eq.
(3.3) has an origin at the heater probe and points downstream.
Assuming amount of released heat, wood density, and wood specic heat capacity are
known, the inferred posterior distributions of the parameters are presented in Figure 4.1,
where the limits of x-axis are the bounds imposed on the parameters as prior distributions,
69
and the true parameter values and their posterior means are represented by the solid and
dashed vertical line, respectively. The results demonstrate that our parameter estimation
process signicantly reduces the parameter uncertainty after conditioning the parameter
values on the observed heat response curves. All the inferred parameters except B
1
are
have tight distributions around their true values. For the estimated thermal diffusivities,
both the mean and mode of the posterior distributions are close to the true values. Whereas
for probe geometries, modes of the posterior distributions are closer to the true values
although the mean value still provides a good estimate. Given that only square terms of
y
u
and y
d
are involved in the analytical models of the heat transport process, y
2
u
and y
2
d
are
estimated rather than y
u
and y
d
.
Each set of sampled parameters from the MCMC method yields an estimation of heat
pulse velocity using Eq. (3.10), and ultimately, a possible model t to the data through
Eq. (3.3). Inclusion of a large number of alternative parameter sets denes the uncertainty
in model tting as well as the estimated heat pulse velocity, which is a key step in eld
applications of the heat ratio method. The distribution of estimated heat pulse velocities
and uncertainty in model tting for the test case are shown in Figure 4.2, from which we
can observe that the model ttings to the data are with a narrow uncertainty range, and the
estimated heat pulse velocities are distributed closely around the true values.
However, it is not surprising that the heat amount, wood density, and wood specic
heat capacity are not known exactly in actual applications, because the heat loss may not
be easily estimated without detailed analysis of the complete circuit and knowledge of how
70
0.01 0 0.01
0
500
1000
B
0
(cm/s)
0.8 1 1.2
0
5
B
1
1 2 3 4 5
x 10
3
0
5000
P
d
f
(cm
2
/s)
1 2 3 4 5
x 10
3
0
5000
(cm
2
/s)
1 2 3 4 5
x 10
3
0
5000
10000
(cm
2
/s)
0.5 1 1.5
0
10
20
30
P
d
f
x
d
(cm)
1.5 1 0.5
0
5
10
15
x
u
(cm)
0 0.5 1
0
10
20
30
y
d
2
(cm
2
)
0 0.5 1
0
5
P
d
f
y
u
2
(cm
2
)
Test1 Test2
Test 3
Figure 4.1: Posterior distributions of parameters with known heat amount, wood density
and wood specic heat capacity. The limits of x-axis represent the bounds imposed on
parameters as prior distributions. The solid vertical lines represent the true parameter values
and dashed vertical lines are the mean values calculated from the posterior distributions.
71
0 20 40 60 80 100 120
0
1
2
3
4
5
Time (s)
T
e
m
p
e
r
a
t
u
r
e
r
i
s
e
(
o
C
)
28 29 30 31 32 33 34 35
0
0.2
0.4
0.6
0.8
Heat pulse velocity (cm/hr)
p
d
f
0 20 40 60 80 100 120
0
1
2
3
4
Time (s)
T
e
m
p
e
r
a
t
u
r
e
r
i
s
e
(
o
C
)
13 14 15 16 17 18 19 20
0
0.2
0.4
0.6
0.8
Heat pulse velocity (cm/hr)
p
d
f
0 20 40 60 80 100 120
0
1
2
3
4
Time (s)
T
e
m
p
e
r
a
t
u
r
e
r
i
s
e
(
o
C
)
8 9 10 11 12 13
0
0.2
0.4
0.6
0.8
Heat pulse velocity (cm/hr)
p
d
f
Figure 4.2: Model tting and statistical distribution of estimated heat pulse velocities for
the test case with known heat amount, wood density, and wood specic heat capacity. In
the left panel, each solid line represents a model t to the data by a parameter set from the
MCMC method. In the right panel, solid vertical lines represent the true heat pulse veloci-
ties and dashed vertical lines are the mean values calculated fromthe posterior distributions.
72
wood density and specic heat capacity change with stem moisture content, which is usu-
ally not measured in situ. In such case, q/(c) in Eq. (3.3) is dened as another unknown
parameter, C, and it can be estimated along with other parameters in our framework.
For the same test case that yield results in Figures 4.1 and 4.2, its counterpart with un-
known C lead to the results in Figures 4.3 and 4.4, where the uncertainty levels in estimated
parameters and heat pulse velocities, as well as in the model tting increase by introducing
uncertainty into C. The results also show that our framework tends to slightly overestimate
C values, which causes overestimation in and v
h
. Nevertheless, in all three tests the mean
values of and v
h
provide reasonable estimations of their underlying true values.
We also studied the case assuming that the true probe geometry is known, in which very
accurate estimates of C, , and v
h
are obtained as shown in Table 4.1. The estimated pa-
rameters are provided as their mean values plus or minus their standard deviations. We can
therefore conclude that, provided with the exact probe geometry, the parameters estimated
by the proposed method converge to the true values with low uncertainty.
It is worth mentioning that the Least-Square estimator is likely to converge to wrong
parameter values when neither C or probe geometry is known, yet provides near perfect
model ts to the data. This observation indicates the existence of multiple solutions under
such conditions. On the other hand, it shows the necessity of using Bayesian technique to
characterize statistical distributions of the parameters rather than an optimized set, which
might be biased.
73
0.01 0 0.01
0
200
400
600
800
B
0
(cm/s)
0.8 1 1.2
0
2
4
6
8
B
1
0.2 0.4 0.6 0.8
0
5
10
P
d
f
C (
C cm
2
)
0.2 0.4 0.6 0.8
0
5
10
15
C (
C cm
2
)
0.2 0.4 0.6 0.8
0
5
10
15
C (
C cm
2
)
1 2 3 4 5
x 10
3
0
1000
2000
3000
P
d
f
(cm
2
/s)
1 2 3 4 5
x 10
3
0
1000
2000
3000
(cm
2
/s)
1 2 3 4 5
x 10
3
0
1000
2000
3000
(cm
2
/s)
0.5 1 1.5
0
5
10
P
d
f
x
d
(cm)
1.5 1 0.5
0
5
10
15
x
u
(cm)
0 0.5 1
0
5
10
y
d
2
(cm
2
)
0 0.5 1
0
5
10
P
d
f
y
u
2
(cm
2
)
Test 1 Test 1 Test 2 Test 3
Test 1 Test 2 Test 3
Figure 4.3: Posterior distributions of parameters with unknown heat amount, wood density
and wood specic heat capacity. The solid lines represent the true parameter values and
dashed vertical lines are the mean values calculated from the posterior distributions.
Table 4.1: Parameters estimated by MCMC method with known probe geometry compared
to true values
Parameter True value Estimated
Test 1
C(
o
C cm
2
) 0.416 0.412 0.006
(10
3
cm
2
/s) 3.0 3.0 0.034
v
h
(cm/hr) 30.0 30.0 0.5
Test 2
C(
o
C cm
2
) 0.416 0.413 0.006
(10
3
cm
2
/s) 3.5 3.5 0.054
v
h
(cm/hr) 15.0 15.0 0.4
Test 3
C(
o
C cm
2
) 0.416 0.412 0.005
(10
3
cm
2
/s) 2.8 2.8 0.036
v
h
(cm/hr) 10.0 10.0 0.4
74
0 20 40 60 80 100 120
0
2
4
Time (s)
T
e
m
p
e
r
a
t
u
r
e
r
i
s
e
(
o
C
)
25 30 35
0
0.2
0.4
Heat pulse velocity (cm/hr)
p
d
f
0 20 40 60 80 100 120
0
2
4
Time (s)
T
e
m
p
e
r
a
t
u
r
e
r
i
s
e
(
o
C
)
12 14 16 18 20 22
0
0.2
0.4
Heat pulse velocity (cm/hr)
p
d
f
0 20 40 60 80 100 120
0
2
4
Time (s)
T
e
m
p
e
r
a
t
u
r
e
r
i
s
e
(
o
C
)
8 10 12 14
0
0.2
0.4
0.6
Heat pulse velocity (cm/hr)
p
d
f
Figure 4.4: Model tting and statistical distributions of estimated heat pulse velocities for
the test case with unknown heat amount, wood density, and wood specic heat capac-
ity. In the left panel, each solid line represents a model t to the data by a parameter set
from the MCMC method. In the right panel, solid vertical lines represent the true heat
pulse velocities and dashed vertical lines are the mean values calculated from the posterior
distributions.
75
4.3.2 Applications to eld data
Our eld tests were conducted on days 221, 234, 251,297, 315 and 329 in year 2007.
When the data were used for parameter estimation, Marshalls model was adopted to t the
heat response curves for its simplicity and computational efciency in repeated sampling
required by the MCMC scheme. We accounted for the signicant discrepancy between
Marshalls idealized model and our more realistic model in calculating early-time temper-
ature increases, by using only the temperature increases measured at times greater than 20s
following the heat pulse. The change in mean and standard deviation of the inferred wood
thermal diffusivity over test period is shown in Figure 4.3, along with the precipitation and
wood temperatures when the tests were conducted. It can be observed that the mean value
of wood thermal diffusivity changes over the dry period, while in wet period, it changes
only slightly. The uncertainty associated with the inferred thermal diffusivity is steady over
the season.
The seasonality of the wood thermal diffusivity can be related to the wood moisture
content, wood temperature, and wood porosity [Steinhagen, 1977; Suleiman et al., 1999;
Simpson and TenWolde, 1999]. In general, wood thermal diffusivity decreases with wood
moisture content and wood porosity while increases with temperature. The example in Ap-
pendix C shows dependence of wood thermal diffusivity on wood moisture content. The
temperature effect is found negligible using the calculation procedure outlined in Appendix
C. It is demonstrated in Figure 4.5 that wood thermal diffusivity changes more signicantly
in the dry period than in the wet period, which might be related to shrinkage of tree trunk
76
3.0
3.2
3.4
3.6
3.8
4.0
T
h
e
r
m
a
l
d
i
f
f
u
s
i
v
i
t
y
(
c
m
2
/
s
)
10
15
20
25
30
W
o
o
d
t
e
m
p
e
r
a
t
u
r
e
(
o
C
)
220 240 260 280 300 320 340
10
20
30
P
r
e
c
i
p
i
t
a
t
i
o
n
(
m
m
)
10
3
Figure 4.5: Seasonal change of inferred wood thermal diffusivity with precipitation and
wood temperature. Means of wood thermal diffusivity are shown in stars and the vertical
lines denote the uncertainty range within one standard deviation. Wood temperature is
shown in circles and precipitation is shown in bars.
77
during the prolonged summer drought. However, our data were limited to late growing sea-
son and winter, tests covering the entire year may better reveal the complex dependence of
wood thermal diffusivity on moisture content and wood temperature. While conducting the
heat response experiment, taking core samples of sapwood or recording tree diameters can
help to relate the seasonal change of wood thermal diffusivity to those factors. Measuring
wood thermal diffusivity on the samples would facilitate model validation as well.
The tting of the Marshalls model with the calibrated parameters to the data of the rst
three tests is shown in Figure 4.6. Evidently, the tting is good despite the discrepancies at
the beginning, which is expected because the Marshalls model tends to overestimate the
temperature increases at the early times.
Also available in Figure 4.6 are the statistical distributions of the estimated heat pulse
velocity associated with each experiment, which show slow ows around 10 cm/hr. Such
direct assessment of uncertainties in measured heat pulse velocity by the proposed frame-
work is of great importance in real applications, because it facilitates further uncertainty
assessment in applications using the sap ow data, which is discussed in detail in next
chapter.
4.4 Conclusions
The heat pulse method is widely used to measure water ux in plants and soil and is
valued for its simple instrumentation and easily automated data collection. However, no
78
0 20 40 60 80 100 120
0
1
2
3
4
Time (s)
T
e
m
p
e
r
a
t
u
r
e
r
i
s
e
(
o
C
)
4 6 8 10 12 14
0
0.1
0.2
0.3
0.4
Heat pulse velocity (cm/hr)
p
d
f
0 20 40 60 80 100 120
1
0
1
2
3
4
Time (s)
T
e
m
p
e
r
a
t
u
r
e
r
i
s
e
(
o
C
)
4 6 8 10 12 14
0
0.1
0.2
0.3
0.4
Heat pulse velocity (cm/hr)
p
d
f
0 20 40 60 80 100 120
1
0
1
2
3
4
Time (s)
T
e
m
p
e
r
a
t
u
r
e
r
i
s
e
(
o
C
)
6 8 10 12 14 16
0
0.1
0.2
0.3
0.4
Heat pulse velocity (cm/hr)
p
d
f
Observed
Fitted
Figure 4.6: Model tting to data and statistical distributions of estimated heat pulse velocity
for the eld test.In the left panel, each solid line represents a model t to the data by a
parameter set from the MCMC method. In the right panel, dashed vertical lines are the
mean values calculated from the posterior distributions.
79
systematic, non-destructive calibration procedure has yet been developed to determine the
site-specic parameters necessary for calculating sap velocity, i.e., wood thermal diffusivity
and probe geometry. In-situ parameter calibration is important in sap ow measurements
since the wood diffusivity can be affected by wood moisture content, wood temperature,
and wood porosity, and the probes can be misaligned during installation due to drilling
difculties. Such parameter calibration is crucial to obtaining the correct transpiration
amount from the sap ow measurements at the plant scale, and consequently affects the
up-scaling of water ux modeling of the soil-vegetation-atmosphere continuum.
In this study, we presented a statistical framework to estimate these parameters from
in-situ heat response curves collected by the implanted probes of heat ratio apparatus. Con-
ditioned on the heat response data, the parameters were inferred using a Bayesian inversion
technique with Markov chain Monte Carlo sampling method. Experiments using synthetic
data showed that multiple tests conducted on the same apparatus can be used to obtain re-
liable statistical distributions of probe geometry as well as wood thermal diffusivity. The
uncertainties in the estimated parameters can be reduced by introducing additional knowl-
edge on heat amount input to the system or probe geometry.
The wood thermal diffusivity is known to be affected wood moisture content, wood
temperature, and wood porosity. Therefore, addressing seasonality of the wood thermal
diffusivity is an important task in sites that experiences contrasting seasons. The parameter
estimation framework proposed in this study can be used to obtain such seasonality by
conducting heat response experiments over different seasons. In our eld site, the wood
80
thermal diffusivity is found to change more signicantly in the dry period than in the wet
period. However, the tests were limited to late growing season and winter, we expect that
the seasonality of the wood thermal diffusivity can be studied thoroughly by conducting
tests over the entire growing season and with complimentary measurements such as taking
core samples of sapwood or recording changes in stem diameter.
The primary advantages of the proposed methodology are threefold: (1) it does not
require known probe geometry or any further intrusive sampling of sapwood, unlike most
other existing work. All the unknown parameters are inverted using the heat response
curves collected by the implanted probes. (2) The Bayesian framework enables direct
quantication of uncertainty in estimated sap ow velocity, which can further facilitate
the uncertainty assessment in water balance study or upscaled water ux involving the sap
ow measurements. (3) Empirical factors are used to relate the heat pulse velocity to the
ratio of temperature increases under asymmetrical probe alignment and these factors were
estimated in this study. With the empirical factors, we were able to model the heat transfer
in sapwood with a simple analytical solution derived on the basis of idealized assumptions,
which ts the observed heat response curves fairly well.
The proposed methodology is ready to be applied to calibrate existing heat ratio sap
ow systems at other sites. It is highly recommended that any study involving sap ow
measurements take temperature response curves routinely to improve the data accuracy. It
is especially useful when alternative transpiration calibration devices such as lysimeters are
not available.
81
Chapter 5
Monitoring and Modeling Water
Dynamics at Plant Scale
5.1 Introduction
Conceptual understanding and mathematical modeling of water dynamics in water-
limited ecosystems have been extensively researched in hydrological and meteorological
studies. As vegetation plays a critical role in uptaking water from soil and transport it
to the atmosphere, many efforts are invested on understanding and modeling the change
of water uptake by plant roots subjected to water stress. Hydrological and meteorological
studies usually adopt different strategies. Many hydrological studies focus on the root-zone
water balance, and the root water uptake is considered as a sink term in water budget [e.g.,
Feddes et al., 1978, 2001; Laio et al., 2001; Guswa et al., 2002; Clemente et al., 1994;
82
Feddes and Raats, 2004; Vrugt et al., 2001]. In these studies, a potential transpiration is
calculated or specied depending on meteorological conditions and vegetation type, then
this amount is distributed to different soil layers based on root density, nally the amount
assigned to each layer is modied in presence of water stress, using a piece-wise linear
reduction factor similar to Figure 2.1. This approach has also been incorporated into nu-
merical models of soil water and solute transport, such as SWAP [van Dam et al., 2008],
and HYDRUS [
Sim unek et al., 2005, 2008].
One drawback of the aforementioned approach in hydrological studies is that the cal-
culation of potential ET does not take into account other environmental stresses, such as
solar radiation and temperature stresses, even though mechanistic model such as Penman-
Monteith (P-M) equation [Monteith and Unsworth, 1990] is sometimes implemented (e.g.,
SWAP model). On the other hand, meteorological studies often assume that plants adjust
their transpiration rates under stressed conditions by means of stomatal conductance, and
stomata respond to various environmental stresses including solar radiation, temperature,
vapor pressure decit, and soil moisture [Jones, 1992]. When P-M equation is applied
to a tree canopy or to a larger scale, all the transpiring and non-transpiration leaves are
treated as if a big leaf, and the bulk conductance is modeled as a function of environmental
stresses, whose effects are assumed to be multiplicative in most empirical studies [Jarvis
and McNaughton, 1986]. Therefore, there is a need to connect the hydrological studies
on soil water dynamics to the meteorological studies on plant transpiration under stressed
condition, such that the soil water stress can be distinguished from the other environmental
83
stresses and the potential plant transpiration can be calculated appropriately.
Another difculty in soil water dynamics modeling is to specify the boundary condi-
tions. The top boundary condition is relatively easier since it is either inltration or soil
evaporation, which can be measured and modeled. However, the bottom boundary condi-
tion is less known due to the difculty of obtaining measurements. It has been a common
practice to assume a free drainage or no ux boundary at bottom [e.g., Laio et al., 2001;
Guswa et al., 2002; Vrugt et al., 2001], and these options are provided in models like
SWAP and HYDRUS. Nevertheless, the boundary conditions may be more complicated in
semi-arid ecosystems as many studies identied groundwater uptake during dry seasons in
these ecosystems [Lubczynski, 2006; Lewis and Burgy, 1964]. The specication of bottom
boundary condition should be considered jointly with the way to distribute total potential
transpiration to different water sources.
One key to understanding the water dynamics in water-limited is to monitor water use
continuously in soil and in plants across different scales. The availability of such data not
only helps to understand the mechanisms, but also provides valuable source for developing
models and validating them. One primary goal of this study is to present a suite of such wa-
ter dynamics data, including sap ow, soil moisture, and other auxiliary data, collected over
the growing season of 2007 on a single tree in a Californian oak-savanna ecosystem. The
concurrent measurements of tree transpiration, root zone soil moisture, and meteorological
forcings are used to identify the the functional relations between the tree transpiration and
environmental stresses, and to estimate the model parameters using statistical tools. The
84
comparison of change in soil water volume and tree transpiration amount provides insights
to the contribution of soil moisture to tree transpiration over the growing season.
5.2 Materials and Methods
5.2.1 Data acquisition
The specic measurements on an individual tree include multiple soil moisture probes
to monitor soil moisture prole vertically and laterally, soil temperature probes for the pur-
pose of energy balance and soil respiration calculations, and sap ow sensors at different
tree heights for measuring tree transpiration. A schematic representation of the experimen-
tal setup is given in Figure 5.1. The soil moisture in root zone is monitored at depths of
5cm, 20cm and 50cm, respectively, and it is monitored at a depth of 20cm along the dripline
and in the open space. Soil temperature is measured at depths of 5cm and 20cm in both the
root zone and the open space.
Sap ow probe sets were manufactured following the instructions given in Burgess
et al. [2001]. We built thermocouple junctions using 36-G copper-constantan wires. They
were put inside micro-capillary tubes lled with silicone heat transfer compound and then
inserted into capped steel hypodermic needles. Each temperature probe consists of two
thermocouple junctions 1.5 cm apart to allow for temperature readings at two radial depths
(1.0 and 2.5 cm) into the sapwood. The same type of temperature probes were also used
to measure the soil temperature in the root zone and in the open space. The heater probes
85
R n
in
Data Logging
System
ECH2O EC-5
Thermocouple
Sap flow sensor
R n
out
R n
tree
=Rn
in
-Rn
out
Figure 5.1: Schematic of measurement equipments installed around each experimental tree
86
were built with coiled 36-G nichrome wire and they were also put inside steel hypodermic
needles. All the needle lengths were 3 cm, and the needle hubs for both the thermocouple
and heater needle bases were sealed with waterproof epoxy resin.
When installing the sap ow sensors, the outer barks were stripped to ensure xylem
contact and a steel drill guide was used to control the distance between probes. The sap
ow sensors were installed on the north side of the tree and were insulated with aluminum
foil to minimize the inuence of diurnal bole temperature uctuation.
Soil moisture at the site has been monitored using ECH
2
O EC-5 sensors (Decagon
Devices, Inc., Pullman, WA). The sensor measures the dielectric constant of the medium
surrounding the probe and outputs a single voltage value, which can be directly related to
volumetric soil water content. All the probes were calibrated in our laboratory using soil
samples taken from the experimental site. After the samples were dried in oven, they were
put into containers with known volume, then different amount of water was added to each
container to obtain a range of volumetric soil water contents in eld. The soil samples
were equilibrated before measurements were taken. For each probe, voltage readings at
each soil water content were recorded and a regression analysis were conducted to relate
voltage readings and real volumetric soil water content. The calibration curves for the ve
sensors involved in this study is shown in Figure 5.2, which show linear relationships.
All the sensors were connected to an AM16/32 multiplexer attached to a CR10X data
logger (Campbell Scientic Inc., Logan, UT). Data were logged every 30 minutes. A
temperature thermistor was attached to the multiplexer for a reference temperature. The
87
200 300 400 500 600 700
0
0.1
0.2
0.3
0.4
0.5
R
2
=0.994
Voltage measured (mV)
V
o
l
u
m
e
t
r
i
c
w
a
t
e
r
c
o
n
t
e
n
t
(
c
m
3
/
c
m
3
)
Data
Regression
200 300 400 500 600 700
0
0.1
0.2
0.3
0.4
0.5
R
2
=0.992
Voltage measured (mV)
200 300 400 500 600 700
0
0.1
0.2
0.3
0.4
0.5
R
2
=0.979
Voltage measured (mV)
200 300 400 500 600 700
0
0.1
0.2
0.3
0.4
0.5
R
2
=0.995
Voltage measured (mV)
V
o
l
u
m
e
t
r
i
c
w
a
t
e
r
c
o
n
t
e
n
t
(
c
m
3
/
c
m
3
)
200 300 400 500 600 700
0
0.1
0.2
0.3
0.4
0.5
R
2
=0.973
Voltage measured (mV)
Figure 5.2: Calibration curves for EC-5 sensors
88
heat pulse was controlled by a relay manufactured at our lab. The program used to collect
all the data is attached in Appendix C.
In addition to the continuous measurements taken on the individual tree, predawn(around
5am) and midday(around 2pm) leaf water potentials were measured on two trees on the site
in the growing seasons of 2007 and 2008. The measurements were taken every one or two
weeks after the last precipitation event in the spring until the trees were dormant. Meteoro-
logical measurements (e.g., solar radiation, air temperature, wind speed, precipitation, etc)
are available from the overstory eddy covariance tower. The net radiation intercepted by
the tree canopy is estimated from the difference between net radiations measured above the
tree canopy (by sensors installed on the eddy covariance tower) and under the tree canopy
(by sensors installed on a 20-m-long railtrack under tree canopies).
The data of water retention characteristics came from three sources. One of them
was estimated from soil texture and bulk density using pedotransfer functions in Rosetta
database (USDA, 1999), which adopts van Genuchten model [van Genuchten, 1980]. A
range of water retention curves was obtained from soil texture and bulk density of 49 sam-
ples collected over a 200 m 200 m area. Another one was from samples analyzed at
the Division of Agriculture and Natural Resources (DANR) Analytical Soils Laboratory,
University of California-Davis. The other set of data was obtained by Liukang Xu in 2002,
using a WP4 Dewpoint PotentiaMeter (Decagon Devices, Inc., Pullman, WA), which mea-
sures water potential of the sample by relating it to the vapor pressure of air in equilibrium
with the sample.
89
5.2.2 Tree transpiration model under water stress
The whole-plant transpiration can be modeled using the following Penman-Monteith
equation [Monteith and Unsworth, 1990] that assumes the tree canopy as a big leaf,
E =
s(R
n
G) +
a
C
p
G
a
VPD
s +(1+G
a
/G
c
)
(5.1)
where is the latent heat of vaporization of water, E is transpiration ux, s is the derivative
of the saturated vapor pressure against temperature, R
n
is the net radiation intercepted by
tree canopy, G is the ground heat ux density,
a
is the air density, C
p
is the specic heat
of air at constant pressure, VPD is the vapor pressure decit at air temperature, is the
psychrometric constant, G
a
is the bulk aerodynamic conductance for heat and water vapor
transfer through the surface layer, and G
c
is the bulk canopy conductance for water vapor
transfer. G
a
can be approximated using friction velocity and wind speed at a reference
height, i.e.,
G
a
=
u
2
(z)
u(z)
(5.2)
where u(z) and u
(z) are wind speed and friction velocity at reference height z, respectively.
The bulk canopy conductance (G
c
) is generally assumed to be affected by the same
factors that control stomatal behavior of a single leaf [Lhomme, 2001]. While the mecha-
nistic modeling of stomatal functioning is extremely difcult, empirical models are often
adopted to relate stomatal conductance to environmental or physiological factors. One of
the most widely used approaches is the Jarvis-type [Jarvis, 1976; Jarvis and McNaughton,
1986; Jones, 1992], which uses a multiplicative form to describe the response of stomata
90
to environmental factors
G
c
= G
cmax
f
1
(VPD) f
2
(T
a
) f
3
(R
g
) f
4
() (5.3)
where G
cmax
represents the maximum canopy conductance when none of the controlling
factors is limiting. Although the selection of controlling factors may vary among sites,
the most extensive collection may be incoming solar radiation (R
g
), vapor pressure decit
(VPD), air temperature (T
a
), CO
2
concentration and soil water status (volumetric soil water
content , soil water potential or leaf water potential) [Lhomme, 2001]. The forms of
controlling functions can be obtained from controlled environmental experiments [Jones,
1992], and some examples of typical functions are available in Jones [1992] and Lhomme
[2001].
The empirical form as shown in 5.3 is not accepted as a priori in this study. Rather,
a statistical approach, alternating conditional expectation (ACE) algorithm [Breiman and
Friedman, 1985], is adopted to identify the optimal functional dependence of bulk canopy
conductance on the environmental factors. ACE identies the optimal transformation of
both dependent and independent variables in multiple regression by maximizing the linear
correlation between the transformed dependent and independent variables. The ACE algo-
rithm was rst adopted in identifying stress functions at a stand level using eddy covariance
measurements by Kiang [2002]. Its primary advantage is that it is totally data-driven and
does not assume parametric functional forms. More details on this method are available
in Appendix E. ACE algorithm enables us to investigate whether a multiplicative form
of stress functions as suggested in 5.3 applies to the tree on which data were collected,
91
and the functional forms can be determined from the transformed variables. Furthermore,
ACE identies if a variable is statistically signicant in the regression after transformation,
which enables us to decide whether to include a variable in the regression equation.
Once the transformations of the variables are determined, the parameters involved can
be estimated through Bayesian inversion technique with MCMC sampling method, as de-
scribed in Chapter 2 and Chapter 4. This framework was again implemented in the WIN-
BUGS software [Lunn et al., 2000]. More details on the Bayesian inference and MCMC
sampling can be found in the Appendix A. Multiple chains of the parameters were simu-
lated in parallel and the modied Gelman-Rubin convergence diagnostic statistics [Brooks
and Gelman, 1998] were used to test the effective convergence of the samples. Each chain
had a sample size of 60,000 parameter sets with the rst 50,000 realizations discarded to
obtain a stationary distribution. One sample out of every twenty was selected to assure
the independence among samples. The resulting pool of samples was used to derive the
joint posterior distribution of the parameters, as well as the marginal distribution of each
parameter.
5.3 Results and Discussions
5.3.1 Meteorological and water status observations
The continuous measurements on the tree started in March of 2007. Soil moisture
data collected since then are presented in Figure 5.3 along with leaf water potential, air
92
temperature, and precipitation observations in 2007 and 2008. Soil moisture data and air
temperature data are averaged values during daytime (8am-6pm). These two years are
relatively dry years with below-average precipitation: 403 mm for 2007 and 257 mm for
the rst 10 months of 2008. Major rainfall events stopped early in 2008 (day 50 compared
to day 100-150 in a normal year). The seasonal pattern of air temperature is representative
for other meteorological variables, such as solar radiation and vapor pressure decit. In
general, the summer is characterized by high solar radiation, high temperature, and high
vapor pressure decit with essentially no precipitation.The results from two typical days
with full leaf foliage during the growing season of 2007 yield an estimation of 50% of net
radiation intercepted by the tree canopy during the course of day, and this value is used
through the growing season.
The dynamics of soil moisture in wet seasons are mainly controlled by the rainfall
events. The magnitude of soil moisture pulse responding to rainfall is damped with depth
as expected. Soil moisture at all depths started to decline rapidly right after the rainfall
stopped, followed by a slowdown of depletion, which may indicate the onset of soil water
stress. In contrast, the decrease in leaf water potential seems to accelerate in summer. This
can be explained by the high non-linearity of soil water retention characteristics when soil
is getting progressively drier, as shown in Figure 5.4. It is found in 2007 that the soil is
wetter in deeper root zone or in places further away from the center of the tree. However,
in the summer of 2008, the soil moisture at 50 cm in the root zone is signicantly drier than
it was in 2007, which might imply change in root water uptake strategy in dry years.
93
0.2
0.3
0.4
0.5
S
W
C
(
c
m
3
/
c
m
3
)
2007 2008
root@5cm
root@20cm
root@50cm
dripline@20cm
open@20cm
0 100 200 300 365
0
1
2
3
4
5
P
r
e
d
a
w
n
L
W
P
(
M
P
a
)
0 100 200 300
0 100 200 300 365
0
10
20
30
D
a
y
t
i
m
e
a
i
r
T
a
i
r
(
o
C
)
Day of year
0 100 200 300 365
50
100
150
200
250
300
P
r
e
c
i
p
i
t
a
t
i
o
n
(
m
m
)
Day of year
air temperature
Figure 5.3: Meteorological and water status observations in 2007 and 2008. Soil moisture
data and air temperature data are averaged values during daytime (8am-6pm). Predawn leaf
water potential (LWP) were averaged from measurements taken at two other trees within
the footprint of the overstory eddy covariance tower.
94
0.00001
0.0001
0.001
0.01
0.1
1
10
100
1000
10000
0 0.2 0.4 0.6 0.8 1
Degree of Saturation
S
u
c
t
i
o
n
P
r
e
s
s
u
r
e
(
M
P
a
)
Rosetta
DANR
Lab 2002
Figure 5.4: Soil water retention curve at experimental site. The range of water retention
curves (in solid lines) were obtained from soil texture and bulk density using pedotrans-
fer functions in Rosetta database (USDA, 1999), which adopts van Genuchten model [van
Genuchten, 1980]. Data points in squares were from samples analyzed at the DANR Ana-
lytical Soils Laboratory, University of California-Davis. Data shown in dots were obtained
by Liukang Xu in 2002, using a WP4 Dewpoint PotentiaMeter.
95
5.3.2 Sap ow observations
The heat pulse velocities were calculated from the ratio of temperature increases fol-
lowing the procedures outlined in the previous chapter, with parameters calibrated using
heat response curves collected on this specic tree. The data points uctuate beyond three
standard deviations from daily mean were regarded random spikes and were removed from
the time series. A snapshot of heat pulse velocities between day 150 and day 160 of 2007
from the bottom set of probes is given in Figure 5.5. The results show that uncertainty in
estimated heat pulse velocities increases with velocity magnitude, and the velocities mea-
sured at outer section are higher than those measured deeper into the trunk. The data also
show non-zero transpiration at night-time, which was also found in another independent
study at the site [Fisher et al., 2007].
The heat pulse velocities were corrected for wounding effect with a linear factor adopted
from Burgess et al. [2001]. The corrected heat pulse velocities were then converted to sap
ux density following Marshall [1958] and Burgess et al. [2001]. The conversion factor is
dependent on wood moisture content, which is assumed to vary from 0.5 in wet season to
0.25 in dry season. A detailed description of wood moisture relations of sap ux calculation
is given in Appendix D.
Sapwood area is an essential parameter to calculate sap uxes from the sap ux density.
A power-law equation is tted on sapwood area and tree diameter based on data collected
from fresh tree samples cut down by the ranch owner occasionally. The boundary between
sapwood and heartwood was identied from the change of color. The data and the regres-
96
150 151 152 153 154 155 156 157 158 159 160
0
10
20
30
40
V
h
(
c
m
/
h
r
)
Inside probe pair
150 151 152 153 154 155 156 157 158 159 160
0
10
20
30
40
Day of year 2007
V
h
(
c
m
/
h
r
)
Outside probe pair
Figure 5.5: Heat pulse velocities measured from the bottom set of sap ow sensors. Inside
and outside probe pairs are 2.5 cm and 1 cm into the tree sapwood, respectively. The
scatters are mean values calculated from MCMC samples and dashed lines are plus minus
two standard deviations from the mean value.
97
10 20 30 40 50 60 70 80
0
100
200
300
400
500
600
700
800
900
1000
S
a
p
w
o
o
d
a
r
e
a
(
c
m
2
)
Diameter (cm)
y=1.32x
1.5
Figure 5.6: Relation between tree diameter and sapwood area. Measurements of diameter
and sapwood area were conducted on fresh tree samples cut down by the ranch owner oc-
casionally. The boundary between sapwood and heartwood was identied from the change
of color.
sion equation are given in Figure 5.6.
The half-hour sap uxes were added up during day-time (8am to 6pm) as the daily
amount. The night-time transpiration was removed from the study because the mechanism
of night-time transpiration is different from that controls day-time transpiration, and P-M
equation is not suitable for the conditions with low available energy. Figure 5.7 shows
the daily transpiration on leaf area basis in the growing season of 2007. An increasing
trend in the early growing season and declining trend in the summer is clearly observed
from the data. The transpiration data also responds well to the leaf-out date, around day
98
80 90 100 110 120 130 140 150 160 170 180 190 200 210 220 230 240 250 250
0
0.5
1
1.5
2
2.5
3
3.5
Day of year 2007
M
e
a
s
u
r
e
d
t
r
a
n
s
p
i
r
a
t
i
o
n
(
m
m
/
d
a
y
)
Figure 5.7: Daily transpiration measured from bottom set of sap ow sensors. The scat-
ters are mean values calculated from MCMC samples and dashed lines are two standard
deviations from the mean value.
83, in 2007. The total mean transpiration from day 80 to day 250 adds up to 250mm.
The uncertainty range is two standard deviations below or above the mean value. The sap
ow measurements on the same tree seem problematic in 2008, which may be due to the
growing wound effect to the tree. Therefore, these data in 2008 are not used in this study.
99
5.3.3 Water balance calculation
The total transpiration amount during day-time is compared to the total change in soil
water volume on daily and weekly basis in Figure 5.8 and Figure 5.9, respectively. The
negative change in soil water volume indicates inltration during rainfall events. It is ob-
served that after large rainfall events, the depletion in soil water is more than the amount of
transpiration, which may be a result of water leakage to the deeper soils and loss through
soil evaporation. However, tree transpires much more water than the depletion of soil water
during the dry season, which is a strong evidence of tree tapping water from deeper soil
and groundwater. This factor must be appropriately accounted for in future soil moisture
dynamics modeling work.
5.3.4 Parameter estimation results for bulk canopy conductance
The data used in this study were those collected in water-stressed period (normally
includes late spring and entire summer) since the goal is to investigate the effects of soil
water stress. Root-density weighted soil moisture was calculated from the vertical prole
of soil moisture in the root zone using Eq. (2.5). The soil moisture at the depth of 20 cm
was averaged from the two measurements at the same depth in root zone and at the dripline.
The exponential model for cumulative distribution of root biomass [Jackson et al., 1996]
was again adopted, P(z)=1-
z
, with z being the depth in centimeters and value is chosen
to be 0.976 for the oak trees. Bulk canopy conductances were inverted from the P-M
equation, and ACE transformation were conducted on the inverted G
c
on net radiation,
100
80 100 120 140 160 180 200 220 240
15
10
5
0
5
10
15
W
a
t
e
r
d
e
p
t
h
(
m
m
)
Change in SWC
Transpiration
80 100 120 140 160 180 200 220 240
0
20
40
P
r
e
c
i
p
i
t
a
t
i
o
n
(
m
m
)
Day of year 2007
Figure 5.8: Daily water balance. The range of negative change in soil water is cut to enlarge
the scale of the plot. The actual values for the two large negative changes are -28mm and
-60mm, respectively.
101
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 20
30
20
10
0
10
20
30
Week starting from April 1 of 2007
W
a
t
e
r
d
e
p
t
h
(
m
m
)
Transpiration
Change in SWC
Figure 5.9: Weekly water balance.
102
air temperature, vapor pressure decit and soil moisture, using free statistical software R
(http://cran.r-project.org/). Effect of CO
2
concentration was omitted since the uctuation
of CO
2
concentration within a year or two may not cause signicant change in stomatal
functioning. The ACE algorithm identied signicant variables to be net radiation, vapor
pressure decit, and soil moisture. The air temperature is not signicant mostly because it
is closely related to vapor pressure decit and therefore its effect can be represented by the
vapor pressure decit.
After trying both original and log-transformed variables in ACE procedure, the one
with log-transformed G
c
and VPD resulted in the highest correlation (R
2
=0.99) between
transformed dependent variable and transformed independent variables. The optimal trans-
formations in this case are shown in Figure 5.10. It can be observed that no further trans-
formation is needed for G
c
and VPD, and a piece-wise linear transformation works well
for the net radiation. As for the soil moisture, the optimal transformation appears to be
non-linear. However, a piece-wise linear function can work well for it since identifying
the non-linear form is not easy and it may not perform better than the piece-wise linear
relation. Therefore, a piece-wise linear function is adopted for the transformations of both
net radiation and soil moisture.
The high R
2
value indicates a strong linear relationship between the transformed G
c
and transformed VPD, R
n
, and SWC. It is also evident in the plot of transformed log(G
C
)
on summation of transformed log(VPD), SWC, and R
n
. These results imply that a multi-
plicative form of stress functions on bulk canopy conductance is applicable at the tree level,
103
2 1 0 1 2
1
0
1
log(Gc)
T
r
a
n
s
f
o
r
m
e
d
l
o
g
(
G
c
)
0.0 0.5 1.0 1.5
0
.
5
0
.
0
0
.
5
log(VPD)
T
r
a
n
s
f
o
r
m
e
d
l
o
g
(
V
P
D
)
0.10 0.12 0.14 0.16 0.18 0.20
2
.
0
1
.
0
0
.
0
0
.
5
1
.
0
SWC(cm
3
cm
3
)
T
r
a
n
s
f
o
r
m
e
d
S
W
C
10 12 14 16 18 20
0
.
2
0
0
.
1
0
0
.
0
0
Rn(MJ m
2
)
T
r
a
n
s
f
o
r
m
e
d
R
n
Figure 5.10: Optimal transformations as determined by ACE. The multiple R
2
=0.99. The
units of G
c
and VPD are mm/s and kPa, respectively.
104
2 1 0 1
1
0
1
Sum transformed independent variables
T
r
a
n
s
f
o
r
m
e
d
d
e
p
e
n
d
e
n
t
v
a
r
i
a
b
l
e
Figure 5.11: Optimal transformation of log(G
c
) vs the sum of the optimal transformations
of log(VPD), SWC, and R
n
. The solid line is the 1:1 line.
although it is found not appropriate at the stand level [Kiang, 2002].
In summary, the following equation can be applied to describe the dependence of G
c
on
VPD, SWC and R
n
,
log(G
c
) = b
0
+k
VPD
log(VPD) +k
) +k
R
(R
n
R
n
). (5.4)
The parameters in Eq. 5.4 are statistically inverted conditioned on day-time tree tran-
spiration measurements deduced from sap ow method, soil moisture measurements, and
105
meteorological measurements. The prior distributions assigned to the parameters are
b
0
uniform(0, 2.2)
k
VPD
uniform(-1, 0.3)
uniform(0.12, 0.15)
k
uniform(2, 100)
R
n
uniform(15, 18)
k
R
uniform(-0.06, 0.005) (5.5)
where the unit of G
cmax
is mm/s and the unit of VPD is kPa. The posterior marginal
distributions of the parameters are given in Figure 5.12. Compared to their uniform prior
distributions, the posterior distributions are bounded by much narrower ranges with well-
dened peak densities, which implies a reduction in uncertainty and the parameters were
successfully inferred with the Bayesian framework.
The mean values of tted transpiration and tted bulk canopy conductance based on
the parameters estimated from the MCMC method are compared to the mean values of
observed transpiration and inverted bulk canopy conductance in Figure 5.13. The scatters
are observed to distribute closely around the 1:1 line, which indicate good model tting.
Further diagnosis plots in Figure 5.14 show that residuals, which are dened as the differ-
ence between the tted values and observed values, appear to be independent on SWC, net
radiation, VPD and air temperature. This suggests a good tting to the model and there is
no other signicant factors left out.
106
1.9 1.95 2 2.05
0
10
20
30
40
50
60
b
0
P
d
f
Posterior
Prior
1 0.98 0.96 0.94 0.92 0.9
0
10
20
30
40
50
60
P
d
f
k
VPD
0.12 0.122 0.124 0.126 0.128 0.13
0
1000
2000
3000
4000
P
d
f
80 78 76 74 72 70
0
0.2
0.4
0.6
0.8
1
P
d
f
k
16 16.5 17 17.5 18 18.5 19
0
0.5
1
1.5
2
2.5
3
P
d
f
R
n
0.05 0.045 0.04 0.035 0.03 0.025 0.02
0
50
100
150
200
P
d
f
k
R
Figure 5.12: Posterior marginal distributions of the G
c
stress function parameters. The
vertical dashed lines are the mean values calculated from the posterior samples.
107
0 0.5 1 1.5 2 2.5
0
0.5
1
1.5
2
2.5
Mean of observed transpiration(mm/day)
M
e
a
n
o
f
f
i
t
t
e
d
t
r
a
n
s
p
i
r
a
t
i
o
n
(
m
m
/
d
a
y
)
0 2 4 6
0
1
2
3
4
5
6
7
Mean of observed G
c
(mm/day)
M
e
a
n
o
f
f
i
t
t
e
d
G
c
(
m
m
/
d
a
y
)
Figure 5.13: Model tting to observed transpiration and bulk canopy conductance. The
solid lines are 1:1 line
108
0 1 2 3 4 5 6
0.4
0.3
0.2
0.1
0
0.1
0.2
0.3
VPD(kPa)
R
e
s
i
d
u
a
l
(
m
m
/
d
a
y
)
0.1 0.15 0.2
0.4
0.3
0.2
0.1
0
0.1
0.2
0.3
R
e
s
i
d
u
a
l
(
m
m
/
d
a
y
)
10 12 14 16 18 20
0.4
0.3
0.2
0.1
0
0.1
0.2
0.3
Rn(MJ/m
2
)
R
e
s
i
d
u
a
l
(
m
m
/
d
a
y
)
15 20 25 30 35 40
0.4
0.3
0.2
0.1
0
0.1
0.2
0.3
Air temperature (
C)
R
e
s
i
d
u
a
l
(
m
m
/
d
a
y
)
Figure 5.14: Diagnosis plots of residuals versus dependent variables.
109
120 140 160 180 200 220 240 260
0
0.5
1
1.5
2
2.5
Day of year 2007
T
r
a
n
s
p
i
r
a
t
i
o
n
(
m
m
/
d
a
y
)
Potential
Actual
Figure 5.15: Potential transpiration calculated in 2007 compared to measured actual
transpiration
With Eq.5.4, the potential tree transpiration can be calculated by setting soil moisture
to be
in Eq. 5.4 and then substitute the G
c
values into the P-M equation. The potential
transpiration calculated for the growing season of 2007 is given in Figure 5.15 for illustra-
tion. It is clearly shown in the plot that soil moisture controls the actual tree transpiration
in the summer time. This calculated potential transpiration can be incorporated in further
soil water dynamics modeling.
110
5.4 Conclusions
Understanding water dynamics under water stress is important for water resources man-
agement in the arid and semi-arid areas. We discussed in this study how plant adjusts tran-
spiration under the stressed conditions, using continuous transpiration and soil moisture
data collected at the plant scale in a Californian oak-savanna. The primary goal of this
study was to connect the soil water dynamics modeling work in hydrological studies to
meteorological studies on tree transpiration responding to environmental stresses including
water stress.
The inuence of soil and atmospheric water stresses on plant-scale transpiration was
investigated through inverting bulk canopy conductance from the Penman-Monteith equa-
tion. The functional dependence of bulk canopy conductance on environmental stresses was
identied using the ACE algorithm, which is a non-parametric data-driven statistical tool.
A multiplicative form of stress functions on tree transpiration is suggested by the data. The
parameters involved in the stress functions were inferred using a Bayesian framework with
the MCMC sampling method. The calibrated model can then be used to estimate the po-
tential transpiration of a tree, which can be incorporated into future soil dynamics models.
The advantages of this procedure are that it accounts for the effects of other environmental
stresses, such as vapor pressure decit and net radiation, on the calculation of potential
transpiration, and the parameters inferred using MCMC enables further quantication of
uncertainties involved in the calculated potential transpiration.
A water budget calculation at the plant scale was conducted by comparing the tree
111
transpiration amount with the change in soil water volume in the root zone. It is found
that most water transpired in summer comes from deeper soil or groundwater. This is a
strong evidence showing the groundwater dependence of the semi-arid ecosystem, and it is
an important fact to be considered in further soil water dynamics modeling by specifying
appropriate bottom boundary conditions in dry seasons.
This study can improve further root-zone soil dynamics modeling in semi-arid ecosys-
tems by providing a prototype to include environmental stresses in mechanistic calculation
of potential tree transpiration rate and by showing evidence of tree tapping water from the
deeper soil and groundwater in dry season. Although this study only included data from a
single tree, the fundamental methodology can be applied to data collected on other single
trees and at larger scales. It is also useful for upscaling tree transpiration and soil water
dynamics to larger scales.
112
Chapter 6
Summary
The purpose of this dissertation work was to improve the conceptual understanding and
mathematical modeling of water transport in the soil-plant-atmospheric continuum under
drought conditions, from a plant scale to a stand scale. The particular focus was on inves-
tigating the regulation of soil moisture on plant water use at both scales, for which models
of water stress function were examined and calibrated using water exchange data acquired
in a Californian oak-savanna ecosystem.
With the recognition that successful model development and calibration relies on eld
data, part of the dissertation work was to acquire water use data at plant scale, in addition
to on-going data acquisition efforts at the study site, which included water uxes and me-
teorological variables measured by the eddy covariance tower and soil moisture measured
within the tower footprint. Root zone soil moisture at the plant scale was monitored at
various depths and tree transpiration was monitored using sap ow measurements. The
113
availability of such continuous data sets an example for acquiring water use data at con-
sistent scales and enables the study on how water dynamics at a plant scale respond to
water stress. Furthermore, a network of such measurements on representative trees within
the ecosystem provides valuable information on upscaling water uxes in heterogeneous
environment.
The amount of transpiration by individual trees were monitored by installing sap ow
sensors in the tree trunks. The sapow velocity was inferred from the ratio of temperature
changes measured downstream and upstream of a heating element following the release of
a heat pulse. The theoretical basis of sap ow measurements using the heat ratio method
was reevaluated in this dissertation. An improved solution to the heat transport process
in sapwood was derived by replacing idealized assumptions in Marshalls classic solution
with more realistic ones. More specically, the sapwood was considered bounded porous
medium, the line source had nite length, which may or may not extend through the entire
sapwood depth, and the heat was released from the line source over a duration. Extensive
comparisons on the difference of calculated temperature elds by Marshalls solution and
the improved solution revealed that most signicant discrepancy occurs around the early
times, whereas the difference is negligible at later times. It is therefore recommended that,
when employing the heat ratio method, the temperature increases should be measured in
late time windows.
Numerical experiments based on the improved analytical solution to the heat transport
process in sapwood discovered that the original fundamental equation of the heat ratio
114
method may result in a signicant bias in estimated heat pulse velocity, if the temperature
probes are not installed symmetrically around the heating element. Nevertheless, simple
modications to the intercept and slope of original fundamental equation can account for
the departure of eld experimental setup from the ideal setup. Such modication can also
be accurately determined by the probe geometry. This revised equation was adopted in this
dissertation to calculate sapow velocities.
The successful application of heat ratio method depends on obtaining reliable estima-
tion of sapwood thermal diffusivity and probe geometry. To assure the quality of the sap
ow measurements acquired from the experimental site, this dissertation study also pro-
vided a systematic, non-destructive, and replicable methodology to determine wood ther-
mal diffusivity and probe geometry for the sap ow measurements using the heat ratio
apparatus. The estimation of these parameters was conditioned on the time series of tem-
perature increases, i.e., temperature response curves, monitored by the downstream and
upstream temperature probes after a heat pulse was released by the central heating probe.
The primary advantage of the methodology is that it relies on the information that can
be obtained using the installed probes without any further disturbance to the tree. It can
also be used to obtain seasonality of wood thermal properties by conducting heat response
experiments over different seasons. The proposed methodology is ready to be applied to
calibrate existing heat ratio sap ow systems at other sites. It is highly recommended that
any study involving sap ow measurements take temperature response curves routinely to
improve the data accuracy. It is especially useful when alternative transpiration calibration
115
devices such as lysimeters are not available.
It was discussed in this dissertation howsoil moisture controls actual evapotranspiration
at the stand scale using multi-year eld observations at the daily time step. The inuence
of soil moisture on actual ET was investigated through the dependence of the ratio between
ET, as measured by the eddy covariance towers, and the potential ET, as approximated by
the Priestley-Taylor equation, on the representative soil moisture. The Feddes Model was
found to be in agreement with the observed patterns of soil moisture effects on ET regard-
less of whether the heterogeneous environment of trees and grasses was homogenized or
treated as the sum of its individual components. However, the parameters of the Feddes
Model varied with time. Grasses have lower
2
and higher
1
during wetter years, whereas
trees have the opposite trend, i.e., they start to control ET under moister soil (higher
2
)
during wetter years so that their ET persists into drier soil conditions (i.e., lower
1
). This
inter-annual variability is primarily driven by variations in the seasonal precipitation dis-
tribution pattern from year to year, as well as the responses of various plant functional
types to changes in soil water availability. Grasses tend to rapidly use up the available wa-
ter resources during the wetter years because they have little control over the evaporation,
whereas trees are able to control water loss by limiting the xylem hydraulic conductance
and by controlling their stomata. This difference in responses can also be a result of change
in ne root distributions due to water availability in early seasons.
The selection of representative soil moisture led to considerable difference in the inter-
annual variability of model parameters. Among the various averaging schemes tried in
116
this study, including (1) the arithmetic average of soil moisture measurements at different
depths obtained at one location, (2) the arithmetic average of the spatially distributed soil
moisture measurements at different depths and (3) the root density weighted average of the
distributed soil moisture measurements at different depths, the last scheme yielded the most
consistent behavior of the Feddes Model over time. The study demonstrated that distributed
sampling of soil moisture is necessary to study the effects of soil moisture availability
on ET in open canopy ecosystems. Furthermore, the soil water availability at different
depths has a different inuence on the total ET depending on the fraction of root biomass
present at the corresponding depths. This difference can be accounted for by using the root
density weighted average of the vertical soil moisture prole to produce representative soil
moisture.
The inuence of soil and atmospheric water stresses on plant-scale transpiration was
investigated through inverting bulk canopy conductance from Penman-Monteith equation.
The functional dependence of bulk canopy conductance on environmental stresses was
identied using a non-parametric data-driven statistical tool, ACE method. Amultiplicative
form of stress functions of tree transpiration was suggested by the data. This form can be
used to separate soil water stress from other environmental stresses so that the potential tree
transpiration can be appropriately calculated. A water budget calculation at the plant scale
found that most water transpired in summer comes from deeper soil or groundwater. This is
a strong evidence showing the groundwater dependence of the semi-arid ecosystem, and it
is an important fact to be considered in further soil water dynamics modeling by specifying
117
appropriate boundary conditions in dry seasons. This part of study demonstrated the im-
portance to connect water dynamics studies in both hydrology and meteorology societies.
Although both communities are interested in water transport from soil to the atmosphere
through plant regulation, hydrology community mostly focuses on the root zone soil water
dynamics under drought conditions, whereas the meteorology community is more focused
on studying the mechanisms of plants adapting to changing climate. It is benecial to look
into the efforts in both sides in order to better understand and model the water transport
process along the soil-plant-atmospheric continuum under drought conditions.
This dissertation also demonstrated the strength of Bayesian inverse modeling tech-
nique in dealing with uncertainties arising from various sources in model calibration. This
parameter estimation methodology provided a systematic tool to quantify the parameter un-
certainties conditioned on eld observations. Adopting MCMC sampling scheme enables
direct assessment of uncertainties in predictions that use the parameters or any functions
of the parameters. The improved ability in model calibration along with the availability
of new observations can improve our understanding of the complex mechanisms related to
water dynamics under drought conditions and thus enables newer model development in
water-controlled ecosystems.
118
Bibliography
Andrieu, C., N. Freitas, A. Doucet, and J. M. (2003), An introduction to MCMC for ma-
chine learning, Machine Learning, 50, 543.
Baldocchi, D. (1997), Flux footprints within and over forest canopies, Boundary-Layer
Meteorology, 85(2), 273292.
Baldocchi, D., and T. Meyers (1998), On using eco-physiological, micrometeorological
and biogeochemical theory to evaluate carbon dioxide, water vapor and trace gas uxes
over vegetation: A perspective, Agricultural and Forest Meteorology, 90(1-2), 125.
Baldocchi, D., et al. (2001), Fluxnet: A new tool to study the temporal and spatial variabil-
ity of ecosystem-scale carbon dioxide, water vapor, and energy ux densities, Bulletin of
the American Meteorological Society, 82(11), 24152434.
Baldocchi, D. D., and L. Xu (2007), what limits evaporation from mediterranean oak wood-
lands - the supply of moisture in the soil, physiological control by plants or the demand
by the atmosphere?, Advances in Water Resources, 30(10), 21132122.
119
Baldocchi, D. D., L. K. Xu, and N. Kiang (2004), How plant functional-type, weather,
seasonal drought, and soil physical properties alter water and energy uxes of an oak-
grass savanna and an annual grassland, Agricultural and Forest Meteorology, 123(1-2),
1339.
Barrett, D. J., T. J. Hatton, J. E. Ash, and M. C. Ball (1995), Evaluation of the heat pulse
velocity technique for measurement of sap ow in rainforest and eucalypt forest species
of south-eastern australia, Plant Cell and Environment, 18(4), 463469, 0140-7791.
Bell, J. L., L. C. Sloan, and M. A. Snyder (2004), Regional changes in extreme climatic
events: A future climate scenario, Journal of Climate, 17(1), 8187, 23.
Blonquist, J. M., S. B. Jones, and D. A. Robinson (2005), Standardizing characterization
of electromagnetic water content sensors: Part 2. evaluation of seven sensing systems,
Vadose Zone Journal, 4(4), 10591069.
Botter, G., A. Porporato, I. Rodriguez-Iturbe, and A. Rinaldo (2007), Basin-scale soil mois-
ture dynamics and the probabilistic characterization of carrier hydrologic ows: Slow,
leaching-prone components of the hydrologic response, Water Resources Research, 43,
W02417, doi:10.1029/2006WR005043.
Breiman, L., and J. Friedman (1985), Estimating optimal transformations for multiple re-
gression and correlation, Journal of the Americal Statistical Association, 80(391), 580
598.
120
Bristow, K. L., G. J. Kluitenberg, and R. Horton (1994), Measurement of soil thermal prop-
erties aith a dual-probe heat-pulse technique, Soil Science Society of America Journal,
58(5), 12881294.
Bristow, K. L., G. J. Kluitenberg, C. J. Goding, and T. S. Fitzgerald (2001), A small multi-
needle probe for measuring soil thermal properties, water content and electrical conduc-
tivity, Computers and Electronics in Agriculture, 31(3), 265280.
Brooks, S. P., and A. Gelman (1998), General methods for monitoring convergence of
iterative simulations, Journal of Computational and Graphical Statistics, 7(4), 434455.
Burgess, S. (2008), Sixth international workshop on xylem sap ow, perth, australia,
november 2006, Plant and Soil, 305(1/2), 144 pp.
Burgess, S. S. O. (2006), Measuring transpiration responses to summer precipitation in a
mediterranean climate: a simple screening tool for identifying plant water-use strategies,
Physiologia Plantarum, 127(3), 404412.
Burgess, S. S. O., M. A. Adams, N. C. Turner, and C. K. Ong (1998), The re-
distribution of soil water by tree root systems, Oecologia, 115(3), 306311, doi:
10.1007/s004420050521.
Burgess, S. S. O., M. A. Adams, and T. M. Bleby (2000), Measurement of sap ow in roots
of woody plants: a commentary, Tree Physiology, 20(13), 909913.
Burgess, S. S. O., M. A. Adams, N. C. Turner, C. R. Beverly, C. K. Ong, A. A. H. Khan,
121
and T. M. Bleby (2001), An improved heat pulse method to measure low and reverse
rates of sap ow in woody plants, Tree Physiology, 21(9), 589598.
Campbell, G. S., C. Calissendorff, and J. H. Williams (1991), Probe for measuring soil
specic heat using a heat-pulse method, Soil Science of America Journal, 55(1), 291
293.
Carslaw, H., and J. Jaeger (1959), Conduction of Heat in Solids, 2nd ed., Clarendon Press,
Oxford.
Casper, B. B., H. J. Schenk, and R. B. Jackson (2003), Dening a plants belowground
zone of inuence, Ecology, 84(9), 23132321.
Cermak, J., M. Deml, and M. Penka (1973), New method of sap ow rate determination in
trees, Biologia Plantarum, 15(3), 171178.
Cermak, J., R. Matyssek, and J. Kucera (1993), Rapid response of large, drought-stressed
beech trees to irrigation, Tree Physiology, 12(3), 281290.
Cermak, J., J. Kucera, and N. Nadezhdina (2004), Sap ow measurements with some ther-
modynamic methods, ow integration within trees and scaling up from sample trees to
entire forest stands, Trees: Structure and Function, 18(5), 529546.
Chandra, S., P. A. Lindsey, and N. L. Bassuk (1994), A gauge to measure the mass ow
rate of water in trees, Plant Cell and Environment, 17(7), 867874.
122
Chen, Q., D. Baldocchi, P. Gong, and T. Dawson (2008a), Modeling radiation and pho-
tosynthesis of a heterogeneous savanna woodland landscape with a hierarchy of model
complexity, Agricultural and Forest Meteorology, 148, 10051020.
Chen, X., Y. Rubin, S. Ma, and D. Baldocchi (2008b), Observations and stochastic mod-
eling of soil moisture control on evapotranspiration in a californian oak savanna, Water
Resources Research, 44, W08409, doi:10.1029/2007WR006646.
Cienciala, E., J. Kucera, and A. Lindroth (1999), Long-term measurements of stand water
uptake in swedish boreal forest, Agricultural and Forest Meteorology, 98-9, 547554,
sp. Iss. SI.
Clark, J. S. (2005), Why environmental scientists are becoming bayesians, Ecology Letters,
8(1), 214.
Clark, J. S., and A. E. Gelfand (2006), A future for models and data in environmental
science, Trends in Ecology & Evolution, 21(7), 375380.
Clemente, R. S., R. De Jong, H. N. Hayhoe, R. W. D., and M. Hares (1994), Testing and
comparison of three unsaturated soil water ow models, Agricultural Water Manage-
ment, 25(2), 135 152, doi:10.1016/0378-3774(94)90041-8.
Cohen, Y., and Y. Li (1996), Validating sap ow measurement in eld-grown sunower and
corn, Journal of Experimental Botany, 47(304), 16991707.
123
Cohen, Y., M. Fuchs, and G. C. Green (1981), Improvement of the heat pulse method for
determining sap ow in trees, Plant Cell and Environment, 4(5), 391397.
Cohen, Y., M. Fuchs, V. Falkenug, and S. Moreshet (1988), Calibrated heat pulse method
for determining water uptake in cotton, Agronomy Journal, 80(3), 398402.
Crainiceanu, C. M., J. R. Stedinger, D. Ruppert, and C. T. Behr (2003), Model-
ing the us national distribution of waterborne pathogen concentrations with appli-
cation to cryptosporidium parvum, Water Resources Research, 39(9), 1235, doi:
10.1029/2002WR001664.
Crosbie, R. S., B. Wilson, J. D. Hughes, and C. McCulloch (2007), The upscaling of tran-
spiration from individual trees to areal transpiration in tree belts, Plant and Soil, 297,
223232.
Dawson, T. E., S. S. O. Burgess, K. P. Tu, R. S. Oliveira, L. S. Santiago, J. B. Fisher,
K. A. Simonin, and A. R. Ambrose (2007), Nighttime transpiration in woody plants
from contrasting ecosystems, Tree Physiology, 27(4), 561575.
de Bruin, H. (1983), A model for the priestley-taylor parameter, Journal of Applied Mete-
orology, 22(4), 572578.
de Bruin, H. A. R., and J. Q. Keijman (1979), Priestley-taylor evaporation model applied to
a large, shallowlake in the netherlands, Journal of Applied Meteorology, 18(7), 898903.
Detto, M., N. Montaldo, J. D. Albertson, M. Mancini, and G. Katul (2006), Soil
124
moisture and vegetation controls on evapotranspiration in a heterogeneous mediter-
ranean ecosystem on sardinia, italy, Water Resources Research, 42(8), W08419, doi:
10.1029/2005WR004693.
Dewar, R. C. (2002), The ball-berry-leuning and tardieu-davies stomatal models: synthesis
and extension within a spatially aggregated picture of guard cell function, Plant Cell and
Environment, 25(11), 13831398.
DOdorico, P., F. Laio, A. Porporato, and I. Rodriguez-Iturbe (2003), Hydrologic controls
on soil carbon and nitrogen cycles. ii. a case study, Advances in Water Resources, 26(1),
5970, doi:10.1016/S0309-1708(02)00095-7.
Duursma, R. A., et al. (2008), Predicting the decline in daily maximum transpiration rate
of two pine stands during drought based on constant minimum leaf water potential and
plant hydraulic conductance, Tree Physiology, 28(2), 265276.
Dye, P. J. (1996), Response of eucalyptus grandis trees to soil water decits, Tree Physiol-
ogy, 16(1-2), 233238.
Eamus, D., A. P. OGrady, and L. Hutley (2000), Dry season conditions determine wet
season water use in the wet-dry tropical savannas of northern australia, Tree Physiology,
20(18), 12191226.
Eastham, J., and S. A. Gray (1998), A preliminary evaluation of the suitability of sap
125
ow sensors for use in scheduling vineyard irrigation, American Journal of Enology and
Viticulture, 49(2), 171176.
Esterling, D. R., G. A. Meehl, C. Parmesan, S. A. Changnon, T. R. Karl, and L. O. Mearns
(2000), Climate extremes: Observations, modeling, and impacts, Science, 289(5487),
20682074, doi:10.1126/science.289.5487.2068.
Feddes, R., and P. Raats (2004), Parameterizing the soil-water-plant root system, in
Unsaturated-zone Modeling - Progress, Challenges and Applications, edited by R. A.
Feddes, G. H. d. Rooij, and J. C. v. Dam, Wageningen UR Frontis Series, pp. 95141,
Kluwer Academic Publishers, Dordrecht, The Netherlands.
Feddes, R., P. Kowalik, , and H. Zaradny (1978), Simulation of eld water use and cropy-
ield: Simulation Monograph, John Wiley & Sons Inc, New York, USA.
Feddes, R. A., et al. (2001), Modeling root water uptake in hydrological and climate mod-
els, Bulletin of the American Meteorological Society, 82(12), 27972809.
Federer, C. A. (1979), Soil-plant-atmosphere model for transpiration and availability of
soil-water, Water Resources Research, 15(3), 555562.
Fernandez, J. E., M. J. Palomo, A. Diaz-Espejo, B. E. Clothier, S. R. Green, I. F. Giron, and
F. Moreno (2001), Heat-pulse measurements of sap ow in olives for automating irriga-
tion: tests, root ow and diagnostics of water stress, Agricultural Water Management,
51(2), 99123, 0378-3774.
126
Fisher, J. B., D. D. Baldocchi, L. Misson, T. E. Dawson, and A. H. Goldstein (2007), What
the towers dont see at night: nocturnal sap ow in trees and shrubs at two ameriux
sites in california, Tree Physiology, 27(4), 597610.
Gelfand, A. E., and A. F. M. Smith (1990), Sampling-based approaches to calculating
marginal densities, Journal of the American Statistical Association, 85(410), 398409.
Gelman, A., J. Carlin, H. Stern, and D. Rubin (1995), Bayesian Data Analysis, Texts in
Statistical Science, CRC Press, Boca Raton, Fla.
Gemand, S., and D. Gemand (1984), Stochastic relaxation, gibbs distributions and the
bayesian restoration of images, IEEE Transactions on Pattern Analysis and Machine
Intelligence, 6, 721741.
Geweke, J. (1989), Bayesian inference in econometric models using monte carlo integra-
tion, Econometrica, 24, 13171399.
Gollan, T., N. C. Turner, and E. D. Schulze (1985), The responses of stomata and leaf gas-
exchange to vapor pressure decits and soil water content.3. in the sclerophyllous woody
species nerium-oleander, Oecologia, 65(3), 356362.
Granier, A. (1985), Une nouvelle methode pour la mesure du ux de seve brute dans le
tronc des arbres, Annales des Sciences Forestieres, 42, 193200.
Granier, A., P. Biron, N. Breda, J. Pontailler, and B. Saugier (1996), Transpiration of
127
trees and forest stands: short and long-term monitoring using sapow methods, Global
Change Biology, 2, 265274.
Green, S., B. Clothier, and B. Jardine (2003), Theory and practical application of heat pulse
to measure sap ow, Agronomy Journal, 95(6), 13711379.
Green, S. R., M. B. Kirkham, and B. E. Clothier (2006), Root uptake and transpiration:
From measurements and models to sustainable irrigation, Agricultural Water Manage-
ment, 86(1-2), 165176.
Gribben, R. (1999), On the heat impulse method for deducing sap ow, Journal of Applied
Mathematics & Decision Sciences, 3(2), 171187.
Guswa, A. J., M. A. Celia, and I. Rodriguez-Iturbe (2002), Models of soil moisture dy-
namics in ecohydrology: A comparative study, Water Resources Research, 38(9), 1166,
doi:10.1029/2001WR000826.
Hastings, W. K. (1970), Monte carlo sampling mehtods using markov chains and their
applications, Biometrika, 87, 97109.
Hikaru, K. (2005), Forest categorization according to dry-canopy evaporation rates in the
growing season: comparison of the priestley-taylor coefcient values from various ob-
servation sites, Hydrological Processes, 19(19), 38733896, doi:10.1002/hyp.5987.
Hopmans, J., and K. Bristow (2002), Current capabilities and future needs of root water
128
and nutrient uptake modeling, in ADVANCES IN AGRONOMY, vol. 77, pp. 103183,
Elsevier.
Hou, Z. S., and Y. Rubin (2005), On minimum relative entropy concepts and prior compat-
ibility issues in vadose zone inverse and forward modeling, Water Resources Research,
41(12), W12425, doi:10.1029/2005WR004082.
Ishikawa, C. M., and C. S. Bledsoe (2000), Seasonal and diurnal patterns of soil water
potential in the rhizosphere of blue oaks: evidence for hydraulic lift, Oecologia, 125(4),
459465, doi:10.1007/s004420000470.
Jackson, R. B., J. Canadell, J. R. Ehleringer, H. A. Mooney, O. E. Sala, and E. D. Schulze
(1996), A global analysis of root distributions for terrestrial biomes, Oecologia, 108(3),
389411.
Jarvis, P. (1976), The interpretation of the variations in leaf water potential and stomatal
conductance found in canopies in the eld, Philosophical Transactions of the Royal So-
ciety, London B, 273, 593610.
Jarvis, P., and K. McNaughton (1986), Stomatal control of transpiration: Scaling up from
leaf to region, pp. 1 49, Academic Press, doi:10.1016/S0065-2504(08)60119-1.
Jones, H. G. (1992), Plants and Microclimate: A Quantitative Approach to Environmental
Plant Physiology, 2nd ed., Cambridge University Press, Cambridge, UK.
Katul, G., R. Leuning, and R. Oren (2003), Relationship between plant hydraulic and
129
biochemical properties derived from a steady-state coupled water and carbon transport
model, Plant Cell and Environment, 26(3), 339350.
Kiang, N. (2002), Savannas and seasonal drought: the landscape-leaf connection through
optimal stomatal control, Ph.D. thesis, University of California, Berkeley, California.
Kim, J., Q. Guo, D. D. Baldocchi, M. Y. Leclerc, L. Xu, and H. P. Schmid (2006), Upscaling
uxes from tower to landscape: Overlaying ux footprints on high-resolution (ikonos)
images of vegetation cover, Agricultural and Forest Meteorology, 136(3-4), 132146.
Kluitenberg, G. J., and J. M. Ham (2004), Improved theory for calculating sap ow with
the heat pulse method, Agricultural and Forest Meteorology, 126(1-2), 169173.
Knapp, A. K., et al. (2002), Rainfall variability, carbon cycling, and plant species diversity
in a mesic grassland, Science, 298(5601), 22022205, doi:10.1126/science.1076347.
Kurz-Besson, C., et al. (2006), Hydraulic lift in cork oak trees in a savannah-type mediter-
ranean ecosystem and its contribution to the local water balance, Plant and Soil, 282(1-
2), 361378.
Lagergren, F., and A. Lindroth (2002), Transpiration response to soil moisture in pine and
spruce trees in sweden, Agricultural and Forest Meteorology, 112(2), 6785.
Laio, F., A. Porporato, L. Ridol, and I. Rodriguez-Iturbe (2001), Plants in water-controlled
ecosystems: active role in hydrologic processes and response to water stress: II. proba-
bilistic soil moisture dynamics, Advances in Water Resources, 24(7), 707723.
130
Lefever, R., and O. Lejeune (1997), On the origin of tiger bush, Bulletin of Mathematical
Biology, 59(2), 263294.
Lejeune, O., P. Courteron, and R. Lefever (1999), Short range co-operativity competing
with long range inhibition explains vegetation patterns, acta oecol, Journal of Interna-
tional Ecology, 20(3), 171183.
Lewis, D. C., and R. H. Burgy (1964), Relationship between aak tree roots and groundwater
in fractured rock as determined by tritium tracing, Journal of Geophysical Research,
69(12), 25792588.
Lhomme, J.-P. (2001), Stomatal control of transpiration: Examination of the jarvis-type
representation of canopy resistance in relation to humidity, Water Resources Research,
37(3), 689699.
Lubczynski, M. (2006), Groundwater uxes in arid and semi-arid environments, in Ground-
water and Ecosystems, p. 225.
Lunn, D., A. Thomas, N. Best, and D. Spiegelhalter (2000), Winbugs a bayesian
modelling framework: concepts, structure, and extensibility, Statistics and Computing,
10:325337., 10, 325337.
Ma, S., D. D. Baldocchi, L. Xu, and T. Hehn (2007), Inter-annual variability in carbon
dioxide exchange of an oak/grass savanna and open grassland in california, Agricultural
and Forest Meteorology, 147(3-4), 157171, doi:10.1016/j.agrformet.2007.07.008.
131
Mahfouf, J. F., C. Ciret, A. Ducharne, P. Irannejad, J. Noilhan, Y. Shao, P. Thornton, Y. Xue,
and Z. L. Yang (1996), Analysis of transpiration results fromthe rice and pilps workshop,
Global and Planetary Change, 13(1-4), 7388.
Marshall, D. C. (1958), Measurement of sap ow in conifers by heat transport, Plant Phys-
iology, 33(6), 385396.
McNaughton, K. G., and T. W. Spriggs (1986), A mixed-layer model for regional evapora-
tion, Boundary-Layer Meteorology, 34(3), 243262.
Metropolis, N., A. W. Rosenbluth, A. H. Teller, and E. Teller (1953), Equations of state
calculations by fast computating machine, Journal of Chemical Physics, 21, 10871091.
Miller, G. R., D. D. Baldocchi, B. E. Law, and M. T. (2007), An analysis of
soil moisture dynamics using multi-year data from a network of micrometeoro-
logical observation sites, Advances in Water Resources, 30(5), 10651081, doi:
10.1016/j.advwatres.2006.10.002.
Milly, P. C. D. (1993), An analytical solution of the stochastic storage problem applicable
to soil water, Water Resources Research, 29(11), 37553758.
Monteith, J., and M. Unsworth (1990), Principles of Environmental Physics, 2nd ed.,
Butterworth-Heinemann, Oxford, UK.
Nadezhdina, N., J. Cermak, J. Gasparek, V. Nadezhdin, and A. Prax (2006), Vertical and
132
horizontal water redistribution in norway spruce (picea abies) roots in the moravian up-
land, Tree Physiology, 26(10), 12771288.
National Research Council (NRC) (2008), Integrating Multiscale Observations of U.S. Wa-
ters, National Academies Press, Washington, D.C.
Oren, R., N. Phillips, B. E. Ewers, D. E. Pataki, and J. P. Megonigal (1999), Sap-ux-
scaled transpiration responses to light, vapor pressure decit, and leaf area reduction in
a ooded taxodium distichum forest, Tree Physiology, 19(6), 337347.
Paruelo, J. M., and O. E. Sala (1995), Water losses in the patagonian steppe - a modeling
approach, Ecology, 76(2), 510520.
Porporato, A., P. DOdorico, F. Laio, and I. Rodriguez-Iturbe (2003), Hydrologic controls
on soil carbon and nitrogen cycles. i. modeling scheme, Advances in Water Resources,
26(1), 4558, doi:10.1016/S0309-1708(02)00094-5.
Porporato, A., E. Daly, and I. Rodriguez-Iturbe (2004), Soil water balance and ecosystem
response to climate change, American Naturalist, 164(5), 625632, 35.
Poyatos, R., P. Llorens, J. Pinol, and C. Rubio (2008), Response of scots pine (pinus
sylvestris l.) and pubescent oak (quercus pubescens willd.) to soil and atmospheric water
decits under mediterranean mountain climate, Annals of Forest Science, 65(3), 306p1
306p13.
133
Priestley, C., and R. J. Taylor (1972), On the assessment of surface heat-ux and evapora-
tion using large-scale parameters, Monthly Weather Review, 100(2), 8192.
Rana, G., and N. Katerji (2000), Measurement and estimation of actual evapotranspiration
in the eld under mediterranean climate: a review, European Journal of Agronomy, 13(2-
3), 125153.
Reis, D. S., and J. R. Stedinger (2005), Bayesian MCMC ood frequency analysis with
historical information, Journal of Hydrology, 313(1-2), 97116, sp. Iss. SI.
Ren, T., G. J. Kluitenberg, and R. Horton (2000), Determining soil water ux and pore
water velocity by a heat pulse technique, Soil Science Society of America Journal, 64(2),
552560.
Rietkerk, M., S. C. Dekker, P. C. de Ruiter, and J. van de Koppel (2004), Self-organized
patchiness and catastrophic shifts in ecosystems, Science, 305(5692), 19261929, doi:
10.1126/science.1101867.
Rodriguez-Iturbe, I. (2000), Ecohydrology: A hydrologic perspective of climate-soil-
vegetation dynamics, Water Resources Research, 36(1), 310.
Rodriguez-Iturbe, I., P. A., F. Laio, and L. Ridol (2001a), Intensive or extensive use of
soil moisture: plant strategies to cope with stochastic water availability, Geophysical
Research Letter, 28(23), 22954498.
134
Rodriguez-Iturbe, I., A. Porporato, F. Laio, and L. Ridol (2001b), Plants in water-
controlled ecosystems: active role in hydrologic processes and response to water stress -
I. scope and general outline, Advances in Water Resources, 24(7), 695705.
Rubin, Y. (2003), Applied Stochastic Hydrogeology, Oxford University Press, Oxford, UK.
Saugier, B., A. Granier, J. Pontailler, E. Dufrene, and D. Baldocchi (1997), Transpiration of
a boreal pine forest measured by branch bag, sap ow and micrometeorological methods,
Tree Physiology, 17, 511519.
Scales, J. A., and R. Snieder (1997), To bayes or not to bayes, Geophysics, 62(4), 1045
1046.
Scholes, R. J., and S. R. Archer (1997), Tree-grass interactions in savannas, Annual Review
of Ecology and Systematics, 28(1), 517544.
Scott, R. L., W. L. Cable, and K. R. Hultine (2008), The ecohydrologic signicance of hy-
draulic redistribution in a semiarid savanna, Water Resources Research, 44(2), W02440,
doi:10.1029/2007WR006149.
Service, R. F. (2004), Water resources: As the West goes dry, Science, 303(5661), 1124
1127, doi:10.1126/science.303.5661.1124.
Settin, T., G. Botter, I. Rodriguez-Iturbe, and A. Rinaldo (2007), Numerical studies on
soil moisture distributions in heterogeneous catchments, Water Resources Research, 43,
W05425, doi:10.1029/2006WR005737.
135
Simpson, W., and A. TenWolde (1999), Physical properties and moisture relations of wood
wood handbook : wood as an engineering material, General technical report FPL GTR-
113, USDA Forest Service, Forest Products Laboratory, Madison, WI.
Sketchley, H. R. (1965), Soil survey of the amador area, California, Tech. rep., Soil Con-
servation Service, Department of Agriculture.
Smith, A. F. M., and G. O. Roberts (1993), Bayesian computation via the gibbs sampler
and related markov chain monte carlo methods, Journal of the Royal Statistical Society,
55(1), 323.
Smith, D. M., and S. J. Allen (1996), Measurement of sap ow in plant stems, Journal of
Experimental Botany, 47(305), 18331844.
Smith, T. J., and L. A. Marshall (2008), Bayesian methods in hydrologic modeling: A
study of recent advancements in markov chain monte carlo techniques, Water Resources
Research, 44, W00B05, doi:10.1029/2007WR006705.
Spittlehouse, D. L., and T. A. Black (1981), Agrowing-season water-balance model applied
to 2 douglas-r stands, Water Resources Research, 17(6), 16511656.
Steinhagen, H. P. (1977), Thermal conductive properties of wood, green or dry, from 40
o
c
to 100
o
c : a literature review, Tech. Rep. FPL-9, USDA Forest Service, Madison, WI.
Suleiman, B. M., J. Larfeldt, L. B., and M. Gustavsson (1999), Thermal conductiv-
136
ity and diffusivity of wood, Wood Science and Technology, 33(6), 465473, doi:
10.1007/s002260050130.
Swanson, R. H. (1994), Signicant historical developments in thermal methods for mea-
suring sap ow in trees, Agricultural and Forest Meteorology, 72(1-2), 113132.
Swanson, R. H., and R. Lee (1966), Measurement of water movement from and through
shrubs and trees, Journal of Forestry, 64(3), 187190, 0022-1201.
Swanson, R. H., and D. W. A. Whiteld (1981), Anumerical analysis of heat pulse velocity:
Theory and practice, Journal of Experimental Botany, 32(126), 221239.
Tang, J., and D. D. Baldocchi (2005), Spatial-temporal variation in soil respiration in an
oak-grass savanna ecosystem in california and its partitioning into autotrophic and het-
erotrophic components, Biogeochemistry, 73(1), 183207.
Teuling, A. J., S. I. Seneviratne, C. Williams, and P. A. Troch (2006), Observed timescales
of evapotranspiration response to soil moisture, Geophysical Research Letters, 33(23),
5.
Tierney, L., and A. Mira (1999), Some adaptive monte carlo methods for bayesian infer-
ence, Statistics in Medicine, 18, 25072515.
Tuzet, A., A. Perrier, and R. Leuning (2003), A coupled model of stomatal conductance,
photosynthesis and transpiration, Plant Cell and Environment, 26(7), 10971116.
137
Tyree, M. T., S. D. Davis, and H. Cochard (1994), Biophysical perspectives of xylem evo-
lution - is there a tradeoff of hydraulic efciency for vulnerability to dysfunction, Iawa
Journal, 15(4), 335360.
van Dam, J. C., P. Groenendijk, R. F. Hendriks, and J. G. Kroes (2008), Advances of
Modeling Water Flow in Variably Saturated Soils with SWAP, Vadose Zone Journal,
7(2), 640653, doi:10.2136/vzj2007.0060.
van Genuchten, M. (1980), A closed-form equation for predicting the hydraulic conductiv-
ity of unsaturated soils, Soil Science Society of America Journal, 44, 892898.
Venables, W., and B. Ripley (2003), Modern Applied Statistics with S, 4th ed., Springer,
Heidelberg, Germany.
Vrugt, J. A., J. W. Hopmans, and J.
Sim unek (2001), Calibration of a two-dimensional root
water uptake model, Soil Sci Soc Am J, 65(4), 10271037.
Sim unek, J., M. T. van Genuchten, and M. Sejna (2005), The HYDRUS-1D software pack-
age for simulating the one-dimensional movement of water, heat, and multiple solutes in
variably-saturated media. Version 3.0, HYDRUS Software Series 1, Tech. rep., Depart-
ment of Environmental Sciences,University of California Riverside, Riverside.
Sim unek, J., M. T. van Genuchten, and M. Sejna (2008), Development and applications
of the HYDRUS and STANMOD software packages and related codes, Vadose Zone
Journal, 7(2), 587600, doi:10.2136/vzj2007.0077.
138
Wan, C., I. Yilmaz, and R. E. Sosebee (2002), Seasonal soil-water availability inuences
snakeweed root dynamics, Journal of Arid Environments, 51(2), 255.
Wang, Q. J., T. E. Ochsner, and R. Horton (2002), Mathematical analysis of heat
pulse signals for soil water ux determination, Water Resources Research, 38(6), doi:
10.1029/2001WR001089, 1091.
West, A. G., K. R. Hultine, J. S. Sperry, S. E. Bush, and J. R. Ehleringer (2008), Tran-
spiration and hydraulic strategies in a pinon-juniper woodland, Ecological Applications,
18(4), 911927.
Whitley, R., M. Zeppel, N. Armstrong, C. Macinnis-Ng, I. Yunusa, and D. Eamus (2008),
A modied jarvis-stewart model for predicting stand-scale transpiration of an australian
native forest, Plant and Soil, 305(1-2), 3547.
Williams, C. A., and J. D. Albertson (2004), Soil moisture controls on canopy-scale water
and carbon uxes in an african savanna, Water Resources Research, 40(9), W09302,
doi:10.1029/2004WR003208.
Williams, C. A., and J. D. Albertson (2005), Contrasting short- and long-timescale effects
of vegetation dynamics on water and carbon uxes in water-limited ecosystems, Water
Resources Research, 41(6), W06005, doi:10.1029/2004WR003750.
Wilson, K. B., P. J. Hanson, P. J. Mulholland, D. D. Baldocchi, and S. D. Wullschleger
(2001), A comparison of methods for determining forest evapotranspiration and its com-
139
ponents: sap-ow, soil water budget, eddy covariance and catchment water balance,
Agricultural and Forest Meteorology, 106(2), 153168.
Woodbury, A. D., and Y. Rubin (2000), A full-bayesian approach to parameter inference
from tracer travel time moments and investigation of scale effects at the cape cod exper-
imental site, Water Resources Research, 36(1), 159171.
Woodbury, A. D., and T. J. Ulrych (1993), Minimumrelative entropy - forward probabilistic
modeling, Water Resources Research, 29(8), 28472860.
Xu, L., and D. Baldocchi (2003), Seasonal trends in photosynthetic parameters and stom-
atal conductance of blue oak (quercus douglasii) under prolonged summer drought and
high temperature, Tree Physiology, 23, 865877.
Yokozawa, M., Y. Kubota, and T. Hara (1998), Effects of competition mode on spatial
pattern dynamics in plant communities, Ecological Modelling, 106(1), 116.
140
Appendix A
Parameter estimation for the Feddes
Model
A.1 Parameter Estimation Method
The basis of our parameter estimation approach is the Bayes Theorem given by:
f
M[D
(m[d
) =
f
D[M
(d
[m) f
M
(m)
f
D
(d
)
=
f
D[M
(d
[m) f
M
(m)
m
f
D[M
(d
[m) f
M
(m)d
p
m
(A.1)
where the boldfaced fonts represent vectors, upper-case letters represent random variables,
and lower-case ones represent realizations of random variables. M is a vector of unknown
parameters with dimension p and d
is a vector or matrix of observed data. f
M
(m) denotes
the prior probability density function (pdf) of M, which reects our knowledge of model
141
parameters before observing d
, f
D[M
(d
[m) is the likelihood of observing d
given model
parameters, and f
M[D
(m[d
) is the posterior pdf of M after the observations are taken
into account. The major advantage of Bayesian parameter estimation is that it enables us
to incorporate the prior knowledge about the model parameters into the learning process
through the prior distributions.
Specically, the parameters involved in this study are m=,
1
,
2
. The observations
are R = R
i
(
i
) with the subscript i being the i-th observation. The representative soil
moisture values are taken as observed inputs.
A.1.1 Principle of Minimum Relative Entropy
The selection of prior distributions is critical in the Bayesian approach [Scales and
Snieder, 1997] because the posterior distribution is dependent on the prior distribution
as evident in Eq. (A.1). Instead of choosing arbitrary prior distributions, we adopted a
systematic approach based on the principle of Minimum Relative Entropy (MRE) [Hou
and Rubin, 2005; Rubin, 2003; Woodbury and Rubin, 2000; Woodbury and Ulrych, 1993],
which states that of all the probabilities that satisfy the given constraints, such as average or
higher-order moments, choose the one that has the highest entropy with respect to a known
prior. Since entropy represents the amount of uncertainty associated with a probability
distribution, the principle of MRE favors distribution that is the most uncommitted or the
least subjective with respect to the constraints.
Assume f
0
M
(m) is an initial estimate of the prior distribution f
M
(m), the principle of
142
MRE obtains the optimal estimation of f
M
(m) by minimizing its negative entropy with
respect to the initial estimate subject to known constraints. Its mathematical form is
min
m
f
M
(m)ln
_
f
M
(m)
f
0
M
(m)
_
d
p
m (A.2)
The optimal solution, f
M
(m), has to satisfy the following constraints,
m
f
M
(m)r
i
(m)d
p
m= r
i
, f ori = 1, , N (A.3)
m
f
M
(m)d
p
m= 1 (A.4)
where r
i
is some known functional forms, such as the moments of the data, with r
i
being
the corresponding prior knowledge, and N is the number of constraints in addition to Eq.
(A.4).
For the parameters engaged in the Feddes Model, only their physical bounds are known
as a priori. Their prior distributions are therefore assigned as uniform between the lower
and upper bounds.
A.1.2 Markov Chain Monte Carlo Method
Given the likelihood of data and the prior distributions of the parameters, the inference
of the posterior distribution based on Eq. (A1) is often not tractable, mainly due to the
143
multidimensional integration in the denominator. Sampling or Monte Carlo method has
been extensively used for exploring posterior distribution, one of which is the well-known
importance sampling. However, importance sampling [Geweke, 1989] is of limited use
in high-dimension case because it requires an importance sampling density that has a tail
at least as heavy as the target distribution [Tierney and Mira, 1999]. Markov chain Monte
Carlo (MCMC) method provides an effective alternative sampling strategy in a wide variety
of problems [Gelfand and Smith, 1990; Tierney and Mira, 1999; Andrieu et al., 2003; Smith
and Roberts, 1993], and it has become a widespread tool for Bayesian inference.
The underlying rationale of the MCMC method is to construct a Markov chain in the
parameter space that converges to a stationary distribution, which is the desired posterior
distribution. The chain is constructed such that the next state is only dependent on the
current state according to a transitional probability. There are two general algorithms for the
MCMC method, Metropolis-Hastings algorithm based on the idea of proposal and rejection
[Metropolis et al., 1953; Hastings, 1970] and Gibbs sampler [Gemand and Gemand, 1984;
Gelfand and Smith, 1990] by breaking the joint distribution into a series of conditional
probabilities that are easier to sample. We adopted the latter algorithm because the full
conditional probabilities can be specied for sampling. The samples generated from the
joint posterior distribution allow direct assessment of uncertainty in predictions, using the
parameters or any functions of the parameters.
144
A.1.3 Inference Using WinBUGS
In this study, the MCMC method was implemented using the software package Win-
BUGS [Lunn et al., 2000]. The statistical inference using WinBUGS is based on prior
distributions and a series of conditional probabilities as shown in the following:
R
i
[
i
, lognormal (
i
,
R
)
i
[
i
,
1
,
2
= max
_
1, min
_
0,
i
1
__
i
[
obs,i
,
normal
_
obs,i
,
1
[
2
uniform(0,
2
)
2
uniform(0, 0.5)
uniform(0.1, 2.0)
log(
R
) uniform(-5.0, 2.0)
log(
) uniform(7.0, 1.0), (A.5)
where R
i
is assumed to be lognormally-distributed for its non-negative restriction, with
mean
i
and standard deviation
R
. The uncertainty of R
i
arises from the measurement
errors in the variables involved in the calculation as well as the model uncertainty to esti-
mate the potential ET. Representative soil moisture is modeled as a random variable for its
errors in measurements, interpolation, and aggregation from point measurements to a rep-
resentative value at the stand scale. The standard deviations of both R
i
and
i
are assigned
145
as hyper-parameters that are parameters of unknown parameters. The connection between
1
and
2
is due to the constraint that
1
cannot exceed
2
.
The uniform prior distributions, f (
2
), f (
1
[
2
), f (), f (
) and f (
R
) were se-
lected based on the principle of MRE and their product constitutes the combined prior of
model parameters and hyper-parameters. It is assumed in the inference that R
i
s are inde-
pendent given
i
and , the likelihood is therefore f
D[M
(d[m) f (R[,
1
,
2
, ,
R
,
) =
N
i=1
LN(
i
,
R
).
146
Appendix B
Regression Relations used to Extend
TDR Measurements in 2003-2005 at
Tonzi Site
147
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
Theta probe measurements
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
015cm: using average of all TDRs
y=0.13+1.19x
data
regression
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.13+1.23x
1530cm: using average of all TDRs
Theta probe measurements
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.10+1.19x
3045cm: using average of all TDRs
Theta probe measurements
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.16+1.45x
4560cm: using average of all TDRs
Theta probe measurements
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
Figure B.1: Regression of average TDR measurements on the Theta probe measurements
in 2003. Theta probe measurements are daily averages calculated from continuous data on
half-hour basis. TDR measurements are averages calculated from all TDRs located in the
site. The regression equations are shown along with the regression lines.
148
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
015cm: average of TDRs under canopy
y=0.14+1.17x
data
regression
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.14+1.12x
1530cm: average of TDRs under canopy
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.11+1.23x
3045cm: average of TDRs under canopy
Theta probe measurements
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.17+1.43x
4560cm: average of TDRs under canopy
Theta probe measurements
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
Figure B.2: Regression of TDR measurements under tree canopies on the Theta probe
measurements in 2003. Theta probe measurements are daily averages calculated from con-
tinuous data on half-hour basis. TDR measurements are averages calculated from TDRs
located under tree canopies. The regression equations are shown along with the regression
lines.
149
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
015cm: average of TDRs at open spaces
y=0.13+1.21x
data
regression
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.13+1.32x
015cm: average of TDRs at open spaces
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.10+1.18x
015cm: average of TDRs at open spaces
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
Theta probe measurements
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.15+1.46x
015cm: average of TDRs at open spaces
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
Theta probe measurements
Figure B.3: Regression of TDR measurements in open spaces on the Theta probe measure-
ments in 2003. Theta probe measurements are daily averages calculated from continuous
data on half-hour basis. TDR measurements are averages calculated from TDRs located in
the open spaces (not covered by tree canopy). The regression equations are shown along
with the regression lines.
150
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
Theta probe measurements
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
015cm: using average of all TDRs
y=0.11+1.28x
data
regression
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.17+1.39x
1530cm: using average of all TDRs
Theta probe measurements
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.11+1.24x
3045cm: using average of all TDRs
Theta probe measurements
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.14+1.29x
4560cm: using average of all TDRs
Theta probe measurements
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
Figure B.4: Regression of average TDR measurements on the Theta probe measurements
in 2004. Theta probe measurements are daily averages calculated from continuous data on
half-hour basis. TDR measurements are averages calculated from all TDRs located in the
site. The regression equations are shown along with the regression lines.
151
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
015cm: average of TDRs under canopy
y=0.13+1.33x
data
regression
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.16+1.26x
1530cm: average of TDRs under canopy
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.11+1.25x
3045cm: average of TDRs under canopy
Theta probe measurements
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.12+1.18x
4560cm: average of TDRs under canopy
Theta probe measurements
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
Figure B.5: Regression of TDR measurements under tree canopies on the Theta probe
measurements in 2004. Theta probe measurements are daily averages calculated from con-
tinuous data on half-hour basis. TDR measurements are averages calculated from TDRs
located under tree canopies. The regression equations are shown along with the regression
lines.
152
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
015cm: average of TDRs at open spaces
y=0.10+1.21x
data
regression
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.16+1.40*x
015cm: average of TDRs at open spaces
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.11+1.23*x
015cm: average of TDRs at open spaces
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
Theta probe measurements
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.15+1.42*x
015cm: average of TDRs at open spaces
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
Theta probe measurements
Figure B.6: Regression of TDR measurements in open spaces on the Theta probe measure-
ments in 2004. Theta probe measurements are daily averages calculated from continuous
data on half-hour basis. TDR measurements are averages calculated from TDRs located in
the open spaces (not covered by tree canopy). The regression equations are shown along
with the regression lines.
153
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
Theta probe measurements
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
015cm: using average of all TDRs
y=0.01+0.83x
data
regression
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.00+0.91x
1530cm: using average of all TDRs
Theta probe measurements
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.01+0.91x
3045cm: using average of all TDRs
Theta probe measurements
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.03+1.11x
4560cm: using average of all TDRs
Theta probe measurements
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
Figure B.7: Regression of average TDR measurements on the Theta probe measurements
in 2005. Theta probe measurements are daily averages calculated from continuous data on
half-hour basis. TDR measurements are averages calculated from all TDRs located in the
site. The regression equations are shown along with the regression lines.
154
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
015cm: average of TDRs at open spaces
y=0.02+0.83x
data
regression
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.01+0.89x
015cm: average of TDRs at open spaces
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.03+0.88x
015cm: average of TDRs at open spaces
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
Theta probe measurements
0 0.1 0.2 0.3 0.4 0.5
0
0.1
0.2
0.3
0.4
0.5
y=0.01+1.09x
015cm: average of TDRs at open spaces
T
D
R
m
e
a
s
u
r
e
m
e
n
t
s
Theta probe measurements
Figure B.8: Regression of TDR measurements in open spaces on the Theta probe measure-
ments in 2005. Theta probe measurements are daily averages calculated from continuous
data on half-hour basis. TDR measurements are averages calculated from TDRs located in
the open spaces (not covered by tree canopy). The regression equations are shown along
with the regression lines.
155
Appendix C
Data Logger Program for Data
Collection
;{CR10X}
; data logger program modified from Josh Fishers code
; for Cluster 1
;AM16/32 is running in 2*32 mode
; Take the measurements every half an hour
*Table 1 Program
01: 1800 Execution Interval (seconds)
; Check the battery voltage
1: Batt Voltage (P10)
1: 1 Loc [ Batt_Volt ]
; Check the circuit everyday
2: If time is (P92)
1: 0 Minutes (Seconds --) into a
2: 1440 Interval (same units as above)
3: 30 Then Do
3: Signature (P19)
1: 2 Loc [ Prog_Sig ]
156
4: End (P95)
; Check the panel temp
6: Temp (107) (P11)
1: 1 Reps
2: 12 SE Channel
3: 3 Excite all reps w/E3
4: 3 Loc [ PTemp_C ]
5: 1.0 Mult
6: 0.0 Offset
; Turn on the multiplexor
7: Do (P86)
1: 41 Set Port 1 High
; Take initial temperatures from the thermocouples
; including sap flow sensors and soil temperature
8: Beginning of Loop (P87)
1: 0 Delay
2: 16 Loop Count
; Clock through the multiplexor
9: Do (P86)
1: 72 Pulse Port 2
; Delay to give enough responding time
10: Excitation with Delay (P22)
1: 1 Ex Channel
2: 0 Delay W/Ex (units = 0.01 sec)
3: 1 Delay After Ex (units = 0.01 sec)
4: 0 mV Excitation
; Each measurement is repeated 10 times
11: Beginning of Loop (P87)
1: 0 Delay
2: 10 Loop Count
12: Thermocouple Temp (DIFF) (P14)
1: 1 Reps
2: 1 2.5 mV Slow Range
157
3: 1 DIFF Channel
4: 1 Type T (Copper-Constantan)
5: 3 Ref Temp (Deg. C) Loc [ PTemp_C ]
6: 65 -- Loc [ tempiT_1 ]
7: 1 Mult
8: 0 Offset
13: End (P95)
; Average 10 TC measurements!
14: Spatial Average (P51)
1: 10 Swath
2: 65 First Loc [ tempiT_1 ]
3: 44 -- Avg Loc [ ainitT_1 ]
15: End (P95)
; skip the empty channels
16: Beginning of Loop (P87)
1: 0 Delay
2: 4 Loop Count
17: Do (P86)
1: 72 Pulse Port 2
18: Excitation with Delay (P22)
1: 1 Ex Channel
2: 0 Delay W/Ex (units = 0.01 sec)
3: 1 Delay After Ex (units = 0.01 sec)
4: 0 mV Excitation
19: End (P95)
; take soil moisture measurements
20: Beginning of Loop (P87)
1: 0 Delay
2: 5 Loop Count
; clock the multiplexor!
21: Do (P86)
1: 72 Pulse Port 2
; Delay
158
22: Excitation with Delay (P22)
1: 1 Ex Channel
2: 0 Delay W/Ex (units = 0.01 sec)
3: 1 Delay After Ex (units = 0.01 sec)
4: 0 mV Excitation
; Measure every EC-5 probe 10 times
23: Beginning of Loop (P87)
1: 0 Delay
2: 10 Loop Count
24: Ex-Del-Diff (P8)
1: 1 Reps
2: 5 2500 mV Slow Range
3: 1 DIFF Channel
4: 1 Excite all reps w/Exchan 1
5: 1 Delay (units 0.01 sec)
6: 2500 mV Excitation
7: 65 -- Loc [ tempiT_1 ]
8: 1.0 Mult
9: 0.0 Offset
25: End (P95)
; Average the 10 soil moisture measurements!
26: Spatial Average (P51)
1: 10 Swath
2: 65 First Loc [ tempiT_1 ]
3: 101 -- Avg Loc [ VWC_1 ]
27: End (P95)
; Turn off the multiplexor!
28: Do (P86)
1: 51 Set Port 1 Low
; Reset the HRdivisor to 0
5: Z=X*F (P37)
1: 77 X Loc [ HRdivisor ]
2: 0 F
3: 77 Z Loc [ HRdivisor ]
159
;Turn on the heater and last for 6 seconds
29: Do (P86)
1: 43 Set Port 3 High
30: Excitation with Delay (P22)
1: 1 Ex Channel
2: 0 Delay W/Ex (units = 0.01 sec)
3: 600 Delay After Ex (units = 0.01 sec)
4: 0 mV Excitation
31: Do (P86)
1: 53 Set Port 3 Low
; Set a timer
32: Timer (P26)
1: 0000 Reset Timer
; Measure the temperatures between 60-100s following the heat pulse
33: Beginning of Loop (P87)
1: 0 Delay
2: 0 Loop Count
34: Timer (P26)
1: 76 Loc [ timer ]
; Start the loop when t>60s
35: IF (X<=>F) (P89)
1: 76 X Loc [ timer ]
2: 3 >=
3: 60 F
4: 30 Then Do
; Exit the loop at 100s following the heat pulse
36: IF (X<=>F) (P89)
1: 76 X Loc [ timer ]
2: 3 >=
3: 100 F
4: 31 Exit Loop if True
;Turn on the multiplexor
37: Do (P86)
1: 41 Set Port 1 High
160
;Measure the temperatures from TCs with sap flow sensors
38: Beginning of Loop (P87)
1: 0 Delay
2: 8 Loop Count
39: Do (P86)
1: 72 Pulse Port 2
40: Excitation with Delay (P22)
1: 1 Ex Channel
2: 0 Delay W/Ex (units = 0.01 sec)
3: 1 Delay After Ex (units = 0.01 sec)
4: 0 mV Excitation
41: Thermocouple Temp (DIFF) (P14)
1: 1 Reps
2: 1 2.5 mV Slow Range
3: 1 DIFF Channel
4: 1 Type T (Copper-Constantan)
5: 3 Ref Temp Loc [ PTemp_C ]
6: 24 -- Loc [ postT_1 ]
7: 1.0 Mult
8: 0.0 Offset
42: End (P95)
; Record how many times temperatures are measured on each TC during 60-100s
43: Z=Z+1 (P32)
1: 77 Z Loc [ HRdivisor ]
44: End (P95)
; Turn off Multiplexor
45: Do (P86)
1: 51 Set Port 1 Low
46: End (P95)
; Output data
47: Do (P86)
161
1: 10 Set Output Flag High (Flag 0)
48: Set Active Storage Area (P80)^30133
1: 1 Final Storage Area 1
2: 100 Array ID
; Time when outputting data
49: Real Time (P77)^26495
1: 1221 Year,Day,Hour/Minute,Seconds (midnight = 2400)
; Battery voltage
50: Average (P71)^15633
1: 1 Reps
2: 1 Loc [ Batt_Volt ]
; Panel Temp
51: Average (P71)^19815
1: 1 Reps
2: 3 Loc [ PTemp_C ]
; Average initial temperature
52: Average (P71)^24723
1: 16 Reps
2: 44 Loc [ ainitT_1 ]
; Average temperature following heat pulse
53: Average (P71)^7966
1: 8 Reps
2: 24 Loc [ postT_1 ]
; Soil moisture data
54: Sample (P70)
1: 5 Reps
2: 101 Loc [ VWC_1 ]
*Table 2 Program
01: 0.0000 Execution Interval (seconds)
*Table 3 Subroutines
End Program
162
Appendix D
Sapwood Moisture Relations of Sap
Flow Measurements
The impact of sapwood moisture content on sap ow measurements are reected by
its inuence on thermal diffusivity of fresh wood matrix and the conversion factor from
heat pulse velocity to sap velocity. This investigation is based on the previous work by
Burgess et al. [2001] and Simpson and TenWolde [1999]. The citations are not repeated in
the discussion hereafter. Some denitions of concepts that are related to this discussion are
listed in Table D.1.
Maximum moisture content can be calculated from specic gravity as
M
max
=
1
G
b
1
1.54
(D.1)
where 1.54 is the specic gravity of wood cell walls. Typical values of G
b
in oaks are
around 0.7. Therefore, the corresponding M
max
is around 0.78.
163
Table D.1: Denitions of some concepts related to wood moisture content
Concepts Symbol Denition
green wood
wood in which cell walls are completely
saturated with water
specic gravity G
b
ovendry weight of wood/green volume of wood
Density of water
moisture content M
weight of fresh wood - ovendry weight of wood
ovendry weight of wood
ber saturation point
moisture content at which only the cell walls are
completely saturated while no water exists in cell
lumens
D.1 Moisture Dependence of Thermal Diffusivity
The thermal diffusivity is dened as the ratio of thermal diffusivity to the product of
specic heat capacity and density as the following:
=
K
gw
gw
c
gw
(D.2)
where K
gw
,
gw
and c
gw
are thermal conductivity, density, and specic heat of green wood,
respectively. Therefore, the effect of wood moisture content on wood thermal diffusivity is
based on its effect on K
gw
,
gw
and c
gw
. The properties of green wood and ovendry wood
are distinguished with subscripts gw and dw, respectively.
Density of green wood can be calculated as
gw
= G
b
(1+M) (D.3)
where the unit of
gw
is g/cm
3
and the density of sap water is assumed same as that of pure
water.
164
The heat capacity of green wood is the combination of the heat capacity of dry wood
and sap water. Heat capacity of dry wood is temperature-dependent as
c
dw
= 0.1031+0.003867T (D.4)
where T is wood temperature in kelvin.
For moisture content above ber saturation point, heat capacity of green wood can be
calculated from
c
gw
=
c
dw
+Mc
s
1+M
. (D.5)
Moreover, when the moisture content falls below the ber saturation point, there is an
additional term to account for extra energy in the wood-water bond, and Eq.(D.5) becomes
c
gw
=
c
dw
+Mc
s
1+M
+M(6.191+0.0236T 1.33M). (D.6)
In Eqs.(D.5) and (D.6), heat capacity of sap is taken as 4.182 J/(g.
o
K).
K
gw
is calculated from the following equations:
K
gw
= K
s
MG
b
+K
dw
(1MG
b
) (D.7)
K
dw
= 0.04182(2120F
v
) (D.8)
where F
v
is the void fraction of wood dened as F
v
= 1 G
b
(0.6494 +M), and heat con-
ductivity of sap water is selected as K
s
=0.5984 W/(m.
o
K).
165
D.2 Moisture Dependence of Conversion from Heat Pulse
Velocity to Sap Flux Density
The conversion from heat pulse velocity to sap ux density is given as
J
s
=
gw
c
gw
s
c
s
v
h
= Fv
h
. (D.9)
For moisture contents above the ber saturation point, the conversion factor F can be
rewritten by substituting Eqs. (D.5) and (D.3) into
gw
and c
gw
, and the following equation
is resulted:
F =
G
b
(c
dw
+Mc
s
)
c
s
(D.10)
where the dependence of F on M is linear.
D.3 An Example
For the blue oaks at our experimental site, we assume that the wood moisture content
may vary between 0.6 to 0.25, then the dependence of thermal diffusivity of green wood
and conversion factor F on wood moisture content is provided in Figure D.1 for three
values of G
b
: 0.6, 0.7 or 0.8. This example demonstrates that thermal diffusivity of green
wood increases with the specic gravity while it decreases with wood moisture content.
Furthermore, the dependence is nonlinear. On the other hand, the conversion factor from
heat pulse velocity to sap ux density linearly increases with both wood moisture content
and wood specic gravity.
166
0.2 0.3 0.4 0.5 0.6 0.7
2.8
3
3.2
3.4
3.6
3.8
x 10
3
T
h
e
r
m
a
l
d
i
f
f
u
s
i
v
i
t
y
(
c
m
2
/
s
)
Wood moisture content
G
b
=0.6
G
b
=0.7
G
b
=0.8
0.2 0.3 0.4 0.5 0.6 0.7
0.35
0.4
0.45
0.5
0.55
0.6
0.65
0.7
C
o
n
v
e
r
s
i
o
n
f
a
c
t
o
r
Wood moisture content
G
b
=0.6
G
b
=0.7
G
b
=0.8
Figure D.1: Dependence of wood thermal diffusivity and conversion factor on wood mois-
ture content
167
Appendix E
Alternating Conditional Expectation
Algorithm
The alternating conditional expectation (ACE) algorithm was introduced by Breiman
and Friedman [1985] to identify the optimal transformations of dependent and indepen-
dent variables in multiple regression that produce the maximum linear correlation between
the transformed dependent and independent variables. Such optimal transformations can
be derived by minimizing the unexplained variance of a linear relationship between the
transformed dependent variable and the sum of transformed independent variables.
Consider a given set of dependent variable Y and independent variables X
1
, ..., X
n
, the
general form of regression equation in ACE algorithm is
0
(Y) =
p
i=1
i
(X
i
) + (E.1)
168
where
0
and
i
represent transformations of the dependent and independent variables,
respectively, and p is the dimension of the independent variables. The ACE algorithm
starts with arbitrary transformations with zero mean and unit variance. The unexplained
variance by the regression is dened as
2
(
0
,
1
, ...,
p
) = E
_
0
(Y)
p
i=1
i
(X
i
)
_
2
(E.2)
subject to the constraint E [
0
(Y)] = 1. The minimization of
2
with respect to
0
(Y) and
i
(X
i
) is accomplished through a series of single-function minimizations, which result in
the following equations:
i
(X
i
) = E
_
0
(Y)
j,=i
j
_
X
j
_
[ X
i
_
(E.3)
0
(Y) =
E
_
p
i=1
i
(X
i
) [ Y
| E
_
p
i=1
i
(X
i
) [ Y
|
(E.4)
The equations E.3 and E.4 represent two fundamental steps involved in the ACE algo-
rithm: conditional expectations and iterative minimization, and therefore, the name alter-
nating conditional expectation. The optimal transformations are obtained after the mini-
mization operations, and the transformed dependent and independent variables are related
as follows:
0
(Y) =
p
i=1
i
(X
i
) +
(E.5)
169
where the minimum regression error,
, is related to the maximum correlation coefcient,
, by
2
= 1
2
.
The optimal ACE transformations are derived on the basis of the given data and do
not require a priori assumptions of the functional forms of the dependent and independent
variables. The ACE algorithm can be used to identify the nonlinearity in the original data
and thus provides a powerful tool for exploratory data analysis.
Das könnte Ihnen auch gefallen
- Modelling Mixed ForestDokument1 SeiteModelling Mixed Forestsita_adhiNoch keine Bewertungen
- Database For Educational Courses On Ecosystem Goods and ServicesDokument37 SeitenDatabase For Educational Courses On Ecosystem Goods and Servicessita_adhiNoch keine Bewertungen
- A 0773 eDokument157 SeitenA 0773 esita_adhiNoch keine Bewertungen
- Drug Interaction2eDokument69 SeitenDrug Interaction2eapi-3745953Noch keine Bewertungen
- BiomassDokument14 SeitenBiomasssita_adhiNoch keine Bewertungen
- A4 - Growth and Yield Models For Uneven-Aged StandsDokument21 SeitenA4 - Growth and Yield Models For Uneven-Aged Standssita_adhiNoch keine Bewertungen
- Wilson Howarth PDFDokument13 SeitenWilson Howarth PDFsita_adhiNoch keine Bewertungen
- Adoption of Agroforestry in The Hills of Nepal - UntungDokument20 SeitenAdoption of Agroforestry in The Hills of Nepal - Untungsita_adhiNoch keine Bewertungen
- How To Perform A MANOVA in SPSSDokument9 SeitenHow To Perform A MANOVA in SPSSsita_adhiNoch keine Bewertungen
- Estimation of Biomass and Carbon Stocks in Plants, Soil and ...Dokument8 SeitenEstimation of Biomass and Carbon Stocks in Plants, Soil and ...sita_adhiNoch keine Bewertungen
- SDM OldGrowthForestsDokument8 SeitenSDM OldGrowthForestssita_adhiNoch keine Bewertungen
- 69240c6fbaad77ec02a33c72c38c44f3Dokument48 Seiten69240c6fbaad77ec02a33c72c38c44f3sita_adhiNoch keine Bewertungen
- A4 - Growth and Yield Models For Uneven-Aged StandsDokument21 SeitenA4 - Growth and Yield Models For Uneven-Aged Standssita_adhiNoch keine Bewertungen
- Individual Tree Models For The Crown Biomass Distribution of Scots PineDokument13 SeitenIndividual Tree Models For The Crown Biomass Distribution of Scots Pinesita_adhiNoch keine Bewertungen
- Mook IDokument291 SeitenMook Isita_adhiNoch keine Bewertungen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (120)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- ChE 204 HW-4 and HW-5 Together, Spring 2014, See Changes!Dokument5 SeitenChE 204 HW-4 and HW-5 Together, Spring 2014, See Changes!Irene Kaye AceroNoch keine Bewertungen
- SimScale Simulation Features Overview PDFDokument30 SeitenSimScale Simulation Features Overview PDFRohith ChakkingalNoch keine Bewertungen
- Heating With Coils and Jackets - Spirax SarcoDokument14 SeitenHeating With Coils and Jackets - Spirax SarcoWEN GIE KONGNoch keine Bewertungen
- Heat and Free Energy of Formation of Deuterium OxideDokument20 SeitenHeat and Free Energy of Formation of Deuterium OxiderifikoNoch keine Bewertungen
- PHY 205 Slide 2Dokument46 SeitenPHY 205 Slide 2Idris Usnan100% (1)
- Charles Richet - Thirty Years of Psychical ResearchDokument664 SeitenCharles Richet - Thirty Years of Psychical ResearchBen Steigmann100% (3)
- Reversed Heat Engines - Refrigeration CycleDokument11 SeitenReversed Heat Engines - Refrigeration CycleRachmatNoch keine Bewertungen
- Zero Energy Buildings and Sustainable Development Implications - A ReviewDokument10 SeitenZero Energy Buildings and Sustainable Development Implications - A ReviewJoao MinhoNoch keine Bewertungen
- Kettle ReboilersDokument6 SeitenKettle ReboilersHow Jie WeiNoch keine Bewertungen
- 2002 Beron Vera y Ripa - Seasonal Salinity Balance in The Gulf of CaliforniaDokument15 Seiten2002 Beron Vera y Ripa - Seasonal Salinity Balance in The Gulf of CaliforniaAlejandro Gonzalez SIlvaNoch keine Bewertungen
- Mechanical Engineering ThermofluidsDokument175 SeitenMechanical Engineering Thermofluidsemad11518100% (1)
- Temperature Cross Possible and Impossible ScenarioDokument5 SeitenTemperature Cross Possible and Impossible ScenarioAhmad FarooqNoch keine Bewertungen
- Thermodynamics ChapterDokument11 SeitenThermodynamics ChapterAbhayNoch keine Bewertungen
- Experiment No.: TITLE HERE: Department of Chemical EngineeringDokument10 SeitenExperiment No.: TITLE HERE: Department of Chemical EngineeringCARLO CASTILLONoch keine Bewertungen
- Design 1 Project 2020 - 2Dokument2 SeitenDesign 1 Project 2020 - 2Shahmi Mat NawiNoch keine Bewertungen
- Propiedades Del A Mezcla Naoh Agua PDFDokument5 SeitenPropiedades Del A Mezcla Naoh Agua PDFLuis Alberto Romero VazquezNoch keine Bewertungen
- Final Report FormatDokument4 SeitenFinal Report FormatBashar Al ZoobaidiNoch keine Bewertungen
- Relief PDFDokument0 SeitenRelief PDFscribpenumakaNoch keine Bewertungen
- Aoa 777 Groundwork Hydraulics Transcript PDFDokument10 SeitenAoa 777 Groundwork Hydraulics Transcript PDFberjarry5938100% (1)
- Advanced Process Design in High Volume Kneader Reactors Using Multiple Feed Ports To Avoid Crust Forming Foaming and Low Heat TransferDokument10 SeitenAdvanced Process Design in High Volume Kneader Reactors Using Multiple Feed Ports To Avoid Crust Forming Foaming and Low Heat TransferRaja WajahatNoch keine Bewertungen
- Thermodynamics - Language StrategyDokument3 SeitenThermodynamics - Language StrategyKerby Jean GumallaoiNoch keine Bewertungen
- COCO Help - Heat ExchangerDokument1 SeiteCOCO Help - Heat ExchangerrvramanNoch keine Bewertungen
- Process Technology Level 2: © University of Teesside 2005Dokument38 SeitenProcess Technology Level 2: © University of Teesside 2005Murad ZareebahNoch keine Bewertungen
- Thermal Energy Q (EDITED)Dokument8 SeitenThermal Energy Q (EDITED)Praphul MalolNoch keine Bewertungen
- Food Processing TechnologyDokument12 SeitenFood Processing TechnologyK.P BHOGAYATANoch keine Bewertungen
- SAJT (Winter 2004) Kumana, p13-23Dokument11 SeitenSAJT (Winter 2004) Kumana, p13-23Jimmy KumanaNoch keine Bewertungen
- Nsep 2023 FQP AnalysisDokument6 SeitenNsep 2023 FQP Analysissudhanshusaini2324Noch keine Bewertungen
- Phy105 Secondlaw Entropy2Dokument36 SeitenPhy105 Secondlaw Entropy2boluwatifeajiboye371Noch keine Bewertungen
- Eloiza. DEPARTMENT OF PURE AND APPLIED CHEMISTRYDokument7 SeitenEloiza. DEPARTMENT OF PURE AND APPLIED CHEMISTRYEloiza PromenteraNoch keine Bewertungen
- Assignment #3 - Ala Baniissa - 8210908Dokument3 SeitenAssignment #3 - Ala Baniissa - 8210908musaNoch keine Bewertungen