Das könnte Ihnen auch gefallen
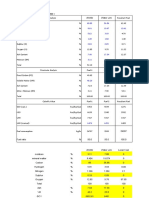
- AreaPercent - VOLATILES CACAO1345837297Dokument2 SeitenAreaPercent - VOLATILES CACAO1345837297Laura NogueraNoch keine Bewertungen
- Qualvolatilescacao 23512Dokument109 SeitenQualvolatilescacao 23512Laura NogueraNoch keine Bewertungen
- Fec2 1Dokument1 SeiteFec2 1Laura NogueraNoch keine Bewertungen
- Food Research International: Luis C. Carrillo, Julián Londoño-Londoño, Andrés GilDokument8 SeitenFood Research International: Luis C. Carrillo, Julián Londoño-Londoño, Andrés GilLaura NogueraNoch keine Bewertungen
- 1 s2.0 S0021967300005355 MainDokument41 Seiten1 s2.0 S0021967300005355 MainLaura NogueraNoch keine Bewertungen
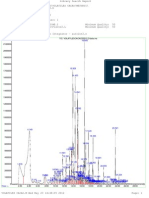
- TIC: FIBRAEXT8512-2.D/data - MS: AbundanceDokument1 SeiteTIC: FIBRAEXT8512-2.D/data - MS: AbundanceLaura NogueraNoch keine Bewertungen
- Croma To GramaDokument1 SeiteCroma To GramaLaura NogueraNoch keine Bewertungen
- Tamaño de GranoDokument7 SeitenTamaño de GranoLaura NogueraNoch keine Bewertungen
- Campos 2010Dokument13 SeitenCampos 2010Laura NogueraNoch keine Bewertungen
- Free Radical Scavenging by Natural Polyphenols: Atom Versus Electron TransferDokument11 SeitenFree Radical Scavenging by Natural Polyphenols: Atom Versus Electron TransferLaura NogueraNoch keine Bewertungen
- Differences Between The Content of Phenolic Compounds in Criollo, Forastero and Trinitario Cocoa Seed (Theobroma Cacao L.)Dokument12 SeitenDifferences Between The Content of Phenolic Compounds in Criollo, Forastero and Trinitario Cocoa Seed (Theobroma Cacao L.)Laura NogueraNoch keine Bewertungen
- Food Research International: Luis C. Carrillo, Julián Londoño-Londoño, Andrés GilDokument8 SeitenFood Research International: Luis C. Carrillo, Julián Londoño-Londoño, Andrés GilLaura NogueraNoch keine Bewertungen
- Effect of Temperature and Air Velocity on Drying Rate and Constant of Cocoa BeansDokument7 SeitenEffect of Temperature and Air Velocity on Drying Rate and Constant of Cocoa BeansLaura NogueraNoch keine Bewertungen
- Textura y Apariencia Del ChocolateDokument19 SeitenTextura y Apariencia Del ChocolateLaura NogueraNoch keine Bewertungen
- Jonfiaessen 2007Dokument5 SeitenJonfiaessen 2007Laura NogueraNoch keine Bewertungen
- Brunetto 2009Dokument5 SeitenBrunetto 2009Laura NogueraNoch keine Bewertungen
- Buffers - Principles and PracticeDokument15 SeitenBuffers - Principles and PracticeLaura NogueraNoch keine Bewertungen
- A Powerful Flavor Impact Constituent of Grapefruit JuiceDokument10 SeitenA Powerful Flavor Impact Constituent of Grapefruit JuiceLaura NogueraNoch keine Bewertungen
- 0 Sensory Evaluation PracticesDokument1 Seite0 Sensory Evaluation PracticesLaura NogueraNoch keine Bewertungen
- Aculey 2010Dokument8 SeitenAculey 2010Laura NogueraNoch keine Bewertungen
- Aculey 2010Dokument8 SeitenAculey 2010Laura NogueraNoch keine Bewertungen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (890)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (587)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (265)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (72)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2219)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (119)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Statutory LPG standardsDokument4 SeitenStatutory LPG standardsalwacsNoch keine Bewertungen
- Arsenic Treatments and CostsDokument284 SeitenArsenic Treatments and CostsG NavarNoch keine Bewertungen
- CabotDokument4 SeitenCabotAliyah AliyNoch keine Bewertungen
- Complex and Protin BindingDokument11 SeitenComplex and Protin BindingdhananjaylandgeNoch keine Bewertungen
- Defend VPS-Putty MSDS 1-1-2011Dokument1 SeiteDefend VPS-Putty MSDS 1-1-2011M. White DentalNoch keine Bewertungen
- Bio 2Dokument12 SeitenBio 2Niyonkuru davidNoch keine Bewertungen
- Chemistry Project - Yap Zhi HengDokument16 SeitenChemistry Project - Yap Zhi HengPuraniNoch keine Bewertungen
- Synthetic Detergents 100 Years of HistoryDokument16 SeitenSynthetic Detergents 100 Years of HistoryJayantha TennakoonNoch keine Bewertungen
- Waste Management in Dyeing UnitsDokument26 SeitenWaste Management in Dyeing UnitsRITA SAHANoch keine Bewertungen
- Caltex Delo Gold SAE 15W40Dokument5 SeitenCaltex Delo Gold SAE 15W40NisarMughalNoch keine Bewertungen
- Notes Chapter 3-StoichiometryDokument31 SeitenNotes Chapter 3-StoichiometryHakim AbbasNoch keine Bewertungen
- ChemicalEngineering Chemical Engineering December 2022Dokument64 SeitenChemicalEngineering Chemical Engineering December 2022Kimberly ConleyNoch keine Bewertungen
- Coal Combustion and Clinker Composition CalculationsDokument5 SeitenCoal Combustion and Clinker Composition CalculationsirfanNoch keine Bewertungen
- Design and Characterization of Nizatidine Effervescent Floating Matrix Tablets Employing Semisynthetic Rate-Retarding PolymersDokument11 SeitenDesign and Characterization of Nizatidine Effervescent Floating Matrix Tablets Employing Semisynthetic Rate-Retarding PolymersSriram NagarajanNoch keine Bewertungen
- BMECME 305 - MSM - Unit 3 - For SharingDokument40 SeitenBMECME 305 - MSM - Unit 3 - For SharingMohamed SohaibNoch keine Bewertungen
- The Effect of Aerophine 3418A Collector Dosage On NiğdeDokument5 SeitenThe Effect of Aerophine 3418A Collector Dosage On NiğdeEduardo CandelaNoch keine Bewertungen
- Beneficiation of Low Grade Graphite Ore of Eastern PDFDokument10 SeitenBeneficiation of Low Grade Graphite Ore of Eastern PDFNarayana Murthy GadiNoch keine Bewertungen
- TOP 10 Co-Extruded Tubes Manufacturers in India.Dokument7 SeitenTOP 10 Co-Extruded Tubes Manufacturers in India.P.Sai Srinivas PrasadNoch keine Bewertungen
- MSDS CpkoDokument4 SeitenMSDS CpkoNicholas Panoguan0% (1)
- 167 Gmaw Zug Asme (Rohmatullah 3g) WPQDokument4 Seiten167 Gmaw Zug Asme (Rohmatullah 3g) WPQMuhammad Fitransyah Syamsuar PutraNoch keine Bewertungen
- Astm A247 10 Standard Test Method For Evaluating The Microstructure of Graphite in Iron Castings - CompressDokument3 SeitenAstm A247 10 Standard Test Method For Evaluating The Microstructure of Graphite in Iron Castings - CompressGigihNoch keine Bewertungen
- Update Question Bank - UNIT VDokument20 SeitenUpdate Question Bank - UNIT VZig zag jeevaNoch keine Bewertungen
- EP2477611B1Dokument2 SeitenEP2477611B1Yahya RizkiNoch keine Bewertungen
- Polymers 13 01943Dokument14 SeitenPolymers 13 01943Visaya L. ClaudineNoch keine Bewertungen
- Demco CatalogDokument50 SeitenDemco CatalogEduardo100% (3)
- Chemistry MCQs Test HandoutsDokument28 SeitenChemistry MCQs Test HandoutsOsama Hasan88% (24)
- Wps PQR WPQ Format - Sa 516 GR 70 2 Zbo7uxDokument5 SeitenWps PQR WPQ Format - Sa 516 GR 70 2 Zbo7uxjobert100% (1)
- Biological Pretreatment of Oil Palm Empty Fruit BunchesDokument1 SeiteBiological Pretreatment of Oil Palm Empty Fruit BunchesIsroi.comNoch keine Bewertungen
- Decomposition of Organic Matter in WaterDokument6 SeitenDecomposition of Organic Matter in WaterDivya Reddy100% (1)
- 3 Unit EMDokument5 Seiten3 Unit EMS.Sampath Chemistry Ghss GurubarapalliNoch keine Bewertungen