Das könnte Ihnen auch gefallen
- GlucosaDokument48 SeitenGlucosaAlejandra Lida Espinoza MormontoyNoch keine Bewertungen
- ORINADokument30 SeitenORINAViridiana MárquezNoch keine Bewertungen
- Patología RenalDokument21 SeitenPatología RenalCarolineReyesAlcántaraNoch keine Bewertungen
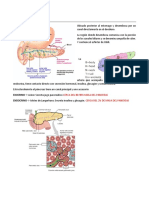
- Anatómica y fisiología del páncreas y hígadoDokument7 SeitenAnatómica y fisiología del páncreas y hígadoGaby Ordoñez0% (1)
- TranscripciónDokument28 SeitenTranscripciónAurelia Gonzales Calle100% (2)
- Gluconeogénesis: síntesis de glucosa a partir de no carbohidratosDokument24 SeitenGluconeogénesis: síntesis de glucosa a partir de no carbohidratosLuis Rodolfo LópezNoch keine Bewertungen
- Crecimiento MicrobianoDokument40 SeitenCrecimiento MicrobianoEduardo RodriguezNoch keine Bewertungen
- Metabolismo de LipidosDokument99 SeitenMetabolismo de LipidosANGUIE ANETTE MAMANI SUCASAIRENoch keine Bewertungen
- Extraccion de Adn A Partir de Sangre Periferica 1Dokument4 SeitenExtraccion de Adn A Partir de Sangre Periferica 1PamelaaGaytan100% (1)
- HematopoyesisDokument19 SeitenHematopoyesisJocelyne Calderon PadillaNoch keine Bewertungen
- Guía endocrinología: conceptos claveDokument14 SeitenGuía endocrinología: conceptos claveluucy0lopez0raNoch keine Bewertungen
- Síntesis de Triglicéridos y Fosfolipidos ANGIEDokument18 SeitenSíntesis de Triglicéridos y Fosfolipidos ANGIEElvis Churacutipa MarcaNoch keine Bewertungen
- OvogenesisDokument15 SeitenOvogenesisRaul CuevaNoch keine Bewertungen
- Pancreas ExocrinoDokument10 SeitenPancreas ExocrinoTamara Alejandra TorresNoch keine Bewertungen
- Sistema Urinario II - HistologíaDokument18 SeitenSistema Urinario II - Histologíabrayan pinedoNoch keine Bewertungen
- La Obesidad - BioquimicaDokument8 SeitenLa Obesidad - BioquimicaGustavo Herrera100% (1)
- Tiroides e HipófisisDokument20 SeitenTiroides e HipófisisJosé ManuelNoch keine Bewertungen
- Importancia Fisiológica de Los AzúcaresDokument4 SeitenImportancia Fisiológica de Los AzúcaresAndrés Báez100% (1)
- Sintesis y Degradacion Del GlucogenoDokument24 SeitenSintesis y Degradacion Del GlucogenoAngel Dominguez RomeroNoch keine Bewertungen
- Liquido AmnioticDokument49 SeitenLiquido AmnioticJhon Andy Ramos100% (1)
- Bioquimica Glucólisis, Gluconeogénesis y GlucógenoDokument14 SeitenBioquimica Glucólisis, Gluconeogénesis y GlucógenoKhris VegaNoch keine Bewertungen
- Órganos linfoides y sus funcionesDokument25 SeitenÓrganos linfoides y sus funcionesviki vargas hernandezNoch keine Bewertungen
- Cap49 LipogénesisDokument20 SeitenCap49 LipogénesisDANIEL ALEJANDRONoch keine Bewertungen
- Circulacion PortalDokument2 SeitenCirculacion PortalEROS ANDREE ZULOETA RAMOSNoch keine Bewertungen
- HEMOGLOBINADokument22 SeitenHEMOGLOBINAapi-3720763100% (1)
- Sistema HematopoyéticoDokument5 SeitenSistema HematopoyéticoChinaw Cueva'sNoch keine Bewertungen
- Mecanismos de Transporte A Través de Membranas BiológicasDokument18 SeitenMecanismos de Transporte A Través de Membranas BiológicasJampieer Sanchez Castillo100% (1)
- Complejo Mayor de HistocompatibilidadDokument5 SeitenComplejo Mayor de HistocompatibilidadFabiana Posligua MolinaNoch keine Bewertungen
- Absorción glucosa glicemiaDokument7 SeitenAbsorción glucosa glicemiaJHERICO YHURAK HUAMAN CERNANoch keine Bewertungen
- Cuadro de ViscerasDokument3 SeitenCuadro de ViscerasDaniel Alfaro100% (1)
- GLANDULA MAMARIA: ESTRUCTURA, IRRIGACIÓN Y DRENAJEDokument3 SeitenGLANDULA MAMARIA: ESTRUCTURA, IRRIGACIÓN Y DRENAJEMarieli Arcos100% (1)
- ComCelMensQuímDokument25 SeitenComCelMensQuímVictoria Sepulveda Salazar100% (1)
- Hormona ParatiroideaDokument8 SeitenHormona ParatiroideagonchigolsNoch keine Bewertungen
- Fisiología del intercambio gaseoso: transporte de O2 y CO2Dokument35 SeitenFisiología del intercambio gaseoso: transporte de O2 y CO2Alejandra de PuertaNoch keine Bewertungen
- GlucogenesisDokument10 SeitenGlucogenesisJavier Martínez LópezNoch keine Bewertungen
- BASOFILOSDokument7 SeitenBASOFILOSHeidy Lisbeth MedinaNoch keine Bewertungen
- Quiz Sistemas Respiratorio y ExcretorDokument6 SeitenQuiz Sistemas Respiratorio y ExcretorJenny Castro JaraNoch keine Bewertungen
- Fisiologia RenalDokument31 SeitenFisiologia RenalKevin Flores100% (7)
- ColiformesDokument22 SeitenColiformesfiorella cipriano0% (1)
- LPS y Receptores Toll (MAOG)Dokument13 SeitenLPS y Receptores Toll (MAOG)miguel angel100% (3)
- Grupos Sanguineos y Sistema HLADokument54 SeitenGrupos Sanguineos y Sistema HLAKari Sdz D100% (1)
- Conversión de La Glucosa 6 Fosfato en GlucosaDokument20 SeitenConversión de La Glucosa 6 Fosfato en GlucosaGabriel RuizNoch keine Bewertungen
- Practica N º 26Dokument23 SeitenPractica N º 26Fernando Junior Parra UchasaraNoch keine Bewertungen
- VolemiaDokument5 SeitenVolemiaAngel Enrique Rodriguez AlvarezNoch keine Bewertungen
- De TodoDokument39 SeitenDe TodoEstela Beatríz Mora GarcíaNoch keine Bewertungen
- Enzimas y cambios energéticosDokument38 SeitenEnzimas y cambios energéticoslisseth carboNoch keine Bewertungen
- Glándulas salivales y sus funcionesDokument55 SeitenGlándulas salivales y sus funcionesDennis Panta DiazNoch keine Bewertungen
- New Grupo SanguineoDokument12 SeitenNew Grupo SanguineoMauricio Caña AvalosNoch keine Bewertungen
- Adipocinas y Sindrome MetabolicoDokument10 SeitenAdipocinas y Sindrome MetabolicoNicole MillanNoch keine Bewertungen
- Actividad Nº8 BIOQUÍMICA PDFDokument18 SeitenActividad Nº8 BIOQUÍMICA PDFZully Denisse Castillo CamaNoch keine Bewertungen
- Grupo SanguineoDokument19 SeitenGrupo SanguineoEdwin Suárez FloresNoch keine Bewertungen
- Sistema AboDokument68 SeitenSistema AboBernardoHernandezNoch keine Bewertungen
- Metabolismo de Los GlúcidosDokument18 SeitenMetabolismo de Los GlúcidosstevemburgNoch keine Bewertungen
- Fisiología de La SangreDokument49 SeitenFisiología de La SangreDaniela Godoy Osorio100% (1)
- Fisiologia RespiratoriaDokument3 SeitenFisiologia RespiratoriaReicitovz JoseNoch keine Bewertungen
- Inflamacion CronicaDokument36 SeitenInflamacion CronicaFrancisco GomezNoch keine Bewertungen
- Clase Tema 6a GlucolisisDokument53 SeitenClase Tema 6a GlucolisisArianaRiosNoch keine Bewertungen
- Resumen Metabolismo de Glúcidos, Regulación Metabolica, Metabolismo Glucogeno y Vias de Las Pentosas FosfatoDokument28 SeitenResumen Metabolismo de Glúcidos, Regulación Metabolica, Metabolismo Glucogeno y Vias de Las Pentosas FosfatocarolinaNoch keine Bewertungen
- UNIDAD IV B - Gluconeoge - Nesis - 21 Abril2016Dokument30 SeitenUNIDAD IV B - Gluconeoge - Nesis - 21 Abril2016Natalia Paz Solar MartinezNoch keine Bewertungen
- 2. GlucólisisDokument90 Seiten2. Glucólisisarely.ibarra8743Noch keine Bewertungen
- Mapa AsmaDokument2 SeitenMapa AsmaDiego PMNoch keine Bewertungen
- Tarea ToxicologiaDokument4 SeitenTarea ToxicologiaDiego PMNoch keine Bewertungen
- FARMACOMODULACIONDokument25 SeitenFARMACOMODULACIONDiego PM100% (1)
- Politica Digesa MinsaDokument34 SeitenPolitica Digesa MinsacrystelzarateNoch keine Bewertungen
- Alimentos Funcionales PDFDokument4 SeitenAlimentos Funcionales PDFDiego PMNoch keine Bewertungen
- HACCPDokument45 SeitenHACCPDiego PMNoch keine Bewertungen
- Alimentos FuncionalesDokument38 SeitenAlimentos FuncionalesDiego PM100% (2)
- Quinolonas PDFDokument7 SeitenQuinolonas PDFDiego PMNoch keine Bewertungen
- SEMANA 1 Salud PublicaDokument28 SeitenSEMANA 1 Salud PublicaDiego PMNoch keine Bewertungen
- Octava Clase Microb. de La Leche, LecheDokument19 SeitenOctava Clase Microb. de La Leche, LecheDiego PMNoch keine Bewertungen
- Exposicion Final Practicas Pre ProfesionalesDokument11 SeitenExposicion Final Practicas Pre ProfesionalesDiego PMNoch keine Bewertungen
- Clase 6Dokument91 SeitenClase 6Diego PMNoch keine Bewertungen
- Especificaciones de Material Medico en UciDokument13 SeitenEspecificaciones de Material Medico en UciDiego PMNoch keine Bewertungen
- Exposicion Practicas Primer NivelDokument14 SeitenExposicion Practicas Primer NivelDiego PMNoch keine Bewertungen
- Quinolonas PDFDokument7 SeitenQuinolonas PDFDiego PMNoch keine Bewertungen
- Anti Tuber Culo SosDokument10 SeitenAnti Tuber Culo SosDiego PMNoch keine Bewertungen
- Acuerdo de Asociacion Transpacifico (TPP)Dokument24 SeitenAcuerdo de Asociacion Transpacifico (TPP)Diego PMNoch keine Bewertungen
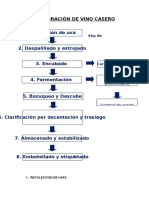
- Elaboración de Vino CaseroDokument7 SeitenElaboración de Vino CaseroDiego PMNoch keine Bewertungen
- Alimentos Funcionales PDFDokument4 SeitenAlimentos Funcionales PDFDiego PMNoch keine Bewertungen
- FARMACOQUÍMICA La Química Medicinal - Ferrufino F, Barrientos R. WWW - Clubdelquimico.tkDokument78 SeitenFARMACOQUÍMICA La Química Medicinal - Ferrufino F, Barrientos R. WWW - Clubdelquimico.tkcyann100% (12)
- Inmunoglobulinas y Otras Moléculas Del Sistema InmuneDokument20 SeitenInmunoglobulinas y Otras Moléculas Del Sistema InmuneDiego PMNoch keine Bewertungen
- Gaussia Luciferase como marcador para monitoreo de tumores metastásicosDokument33 SeitenGaussia Luciferase como marcador para monitoreo de tumores metastásicosDiego PMNoch keine Bewertungen
- Reacciones Adversas MedicamentosasDokument4 SeitenReacciones Adversas MedicamentosasDiego PMNoch keine Bewertungen
- Sistema InmuneDokument11 SeitenSistema InmuneDiego PMNoch keine Bewertungen
- Celulas y OrganosDokument21 SeitenCelulas y OrganosDiego PMNoch keine Bewertungen
- Inhibidores de La Biosisntesis de Acido TetrahidrofolicoDokument25 SeitenInhibidores de La Biosisntesis de Acido TetrahidrofolicoMelyFiallos100% (1)
- Práctica 5 - E6 - 3FM2Dokument7 SeitenPráctica 5 - E6 - 3FM2Fátima Vázquez MonteroNoch keine Bewertungen
- 1 Clase - Tecnologia de Los Alimentos IIIDokument65 Seiten1 Clase - Tecnologia de Los Alimentos IIIElmer Lozada DelgadoNoch keine Bewertungen
- Propiedades Fisicas y Quimicas de Los AlcoholesDokument5 SeitenPropiedades Fisicas y Quimicas de Los AlcoholesDany Angeles0% (2)
- AlquenosDokument13 SeitenAlquenosAlex Daniel ReyesNoch keine Bewertungen
- GLUCOLOSISDokument8 SeitenGLUCOLOSISRoy Michael Guerra ArandaNoch keine Bewertungen
- DANNA MARGARITA SUAREZ DIAZ - 11° Taller Alquenos y Alquinos 2021Dokument15 SeitenDANNA MARGARITA SUAREZ DIAZ - 11° Taller Alquenos y Alquinos 2021DANNA MARGARITA SUAREZ DIAZNoch keine Bewertungen
- Tabla Nutricional ESTANDARDokument22 SeitenTabla Nutricional ESTANDARvictoria valenciaNoch keine Bewertungen
- Clasificacion de AntibioticosDokument3 SeitenClasificacion de Antibioticoshorck 09100% (1)
- Propiedades de las proteínas: solubilidad, desnaturalización y especificidadDokument3 SeitenPropiedades de las proteínas: solubilidad, desnaturalización y especificidadjusellyNoch keine Bewertungen
- Catabolismo de AminoacidosDokument20 SeitenCatabolismo de AminoacidosXimena Véliz DaudNoch keine Bewertungen
- Enzimas - B06 - Andrea Gpe. de La Cruz S.Dokument13 SeitenEnzimas - B06 - Andrea Gpe. de La Cruz S.Andrea SantosNoch keine Bewertungen
- EVALUACION de Quimica Alcanos Alquenos y AlquinosDokument2 SeitenEVALUACION de Quimica Alcanos Alquenos y Alquinosvicmath71% (7)
- Complejo multienzimático de la biosíntesis de ácidos grasosDokument3 SeitenComplejo multienzimático de la biosíntesis de ácidos grasosMichaelRvNoch keine Bewertungen
- Viii Compuestos Carbonilicos II Acidos y DerivDokument36 SeitenViii Compuestos Carbonilicos II Acidos y DerivSarah Duran QuezadaNoch keine Bewertungen
- AminoácidoDokument7 SeitenAminoácidoBetty LopezNoch keine Bewertungen
- Autoevaluación - Integración Metabólica Intento IDokument4 SeitenAutoevaluación - Integración Metabólica Intento IMaría Virginia AlvarezNoch keine Bewertungen
- Guia de Prac N 3 Carbohidratos 2Dokument7 SeitenGuia de Prac N 3 Carbohidratos 2Emerson Maucaylle palominoNoch keine Bewertungen
- Tarea 2 Seres VivosDokument11 SeitenTarea 2 Seres VivosAdriana Del PilarNoch keine Bewertungen
- Alcaloides Derivados Del AATriptófano Version FinalDokument56 SeitenAlcaloides Derivados Del AATriptófano Version FinalMANUELA RAMIREZ SEPULVEDANoch keine Bewertungen
- Receptores MetabotrópicosDokument30 SeitenReceptores MetabotrópicosDorian Mieux PiafNoch keine Bewertungen
- Ejercicio Clase Asincrònica 1Dokument1 SeiteEjercicio Clase Asincrònica 1Steven OrtizNoch keine Bewertungen
- L. Carnitina Como Suplemento NutricionalDokument10 SeitenL. Carnitina Como Suplemento NutricionalGara DigeowenNoch keine Bewertungen
- BMP Ficha Tecnica JurelDokument11 SeitenBMP Ficha Tecnica JurelMarco Godoy AstorgaNoch keine Bewertungen
- Grupos funcionales: ácidos carboxílicos, aldehídos y cetonasDokument12 SeitenGrupos funcionales: ácidos carboxílicos, aldehídos y cetonasFabian SaffontNoch keine Bewertungen
- Ecaes QUIMICA ORGANICA - RESPUESTAS EXAMEN FINALDokument3 SeitenEcaes QUIMICA ORGANICA - RESPUESTAS EXAMEN FINALJorge Roque Villamizar SanchezNoch keine Bewertungen
- Acuerdo 029 2012 RonaldDokument528 SeitenAcuerdo 029 2012 Ronaldbelth almanzaNoch keine Bewertungen
- Propiedades de Alquenos y AlquinosDokument8 SeitenPropiedades de Alquenos y AlquinosLizzyNoch keine Bewertungen
- CoenzimasDokument48 SeitenCoenzimasEduardo HernándezNoch keine Bewertungen
- Informe de CarbohidratosDokument18 SeitenInforme de CarbohidratosAlba ChacónNoch keine Bewertungen