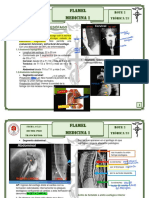
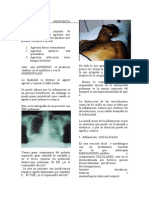
Das könnte Ihnen auch gefallen
- Dime Que Comes y Te Dire Que Bacterias Tienes - 8-20Dokument14 SeitenDime Que Comes y Te Dire Que Bacterias Tienes - 8-20Peter Parker100% (2)
- Glaucoma y sus tiposDokument5 SeitenGlaucoma y sus tiposBianca ArgüellesNoch keine Bewertungen
- Practica 1. FISIODokument5 SeitenPractica 1. FISIOYesleni OrozcoNoch keine Bewertungen
- Protocolo Intoxicacion Por Organofosforados 2013Dokument6 SeitenProtocolo Intoxicacion Por Organofosforados 2013Alexsandra G. VásquezNoch keine Bewertungen
- Tejidos BlandosDokument9 SeitenTejidos Blandossouljazz212Noch keine Bewertungen
- Radio Unido - 230421 - 065520-99-140 PDFDokument42 SeitenRadio Unido - 230421 - 065520-99-140 PDFLuz Betty Alejo MamaniNoch keine Bewertungen
- DCP-sintomas-diagnosticoDokument8 SeitenDCP-sintomas-diagnosticoAlbaNoch keine Bewertungen
- Ontogenia Linfocitos BDokument57 SeitenOntogenia Linfocitos BXimena Granados MuñozNoch keine Bewertungen
- Absceso PulmonarDokument33 SeitenAbsceso Pulmonarsantoslopez100% (1)
- Obtencion Del Dna de BazoDokument13 SeitenObtencion Del Dna de Bazoבריגייט ארטיהNoch keine Bewertungen
- Receptores Citosólicos para Pamp y DampDokument4 SeitenReceptores Citosólicos para Pamp y DampPame FloresNoch keine Bewertungen
- Mecanismos de Evasion de La Respuesta InmuneDokument8 SeitenMecanismos de Evasion de La Respuesta InmuneNnznznznNoch keine Bewertungen
- Capítulo 4 Obstetricia de WilliamsDokument24 SeitenCapítulo 4 Obstetricia de WilliamsAlejandra Vera AlvarezNoch keine Bewertungen
- DIABETES MELLITUS-usmpDokument13 SeitenDIABETES MELLITUS-usmpCristhian Jeanpierre Cueva AlvaresNoch keine Bewertungen
- Muerte Celular y ApoptosisDokument12 SeitenMuerte Celular y ApoptosisCindy PcNoch keine Bewertungen
- Vías LagrimalesDokument21 SeitenVías LagrimalesAngie EncarnacionNoch keine Bewertungen
- Shock AnafilácticoDokument9 SeitenShock AnafilácticoOscar Jativa BravoNoch keine Bewertungen
- Sensibilidad Somatica 3 1222790501483926 8Dokument29 SeitenSensibilidad Somatica 3 1222790501483926 8ajseals0% (1)
- Reflejos FisiologiaDokument5 SeitenReflejos FisiologiaYaniris Sofia100% (1)
- Señalizacion CelularDokument12 SeitenSeñalizacion CelularYean Paul Cruz SuarezNoch keine Bewertungen
- La CardiolipinaDokument10 SeitenLa Cardiolipinauniversal04-82100% (1)
- Mapa Conceptual MorfofisiologiaDokument10 SeitenMapa Conceptual MorfofisiologiaLuzYinethOspinaTovarNoch keine Bewertungen
- Capítulo 6. Técnicas Diagnósticas en HistopatologíaDokument11 SeitenCapítulo 6. Técnicas Diagnósticas en HistopatologíaRaúl Martinez SilvaNoch keine Bewertungen
- Autoevaluacion 2 PDFDokument4 SeitenAutoevaluacion 2 PDFBS3091Noch keine Bewertungen
- Reporte de Un Caso Clinico Una Lesión Relacionada Con El Virus Papiloma HumanoDokument3 SeitenReporte de Un Caso Clinico Una Lesión Relacionada Con El Virus Papiloma HumanoJezzy ContrerasNoch keine Bewertungen
- La SalivaDokument6 SeitenLa SalivaBety Jim AraNoch keine Bewertungen
- Patología orgánica y diagnósticosDokument28 SeitenPatología orgánica y diagnósticosMariela Mina RiveraNoch keine Bewertungen
- Agentes Carcinogenos, Manifestaciones Clinicas, Defensas, Diagnostico.Dokument55 SeitenAgentes Carcinogenos, Manifestaciones Clinicas, Defensas, Diagnostico.Osorio Galindo Kenny Osorio100% (1)
- Psicologia MedicaDokument40 SeitenPsicologia MedicaTommy SalgueiroNoch keine Bewertungen
- Examen físico ocular: Exploración del fondo de ojoDokument5 SeitenExamen físico ocular: Exploración del fondo de ojoCarlos RasmijnNoch keine Bewertungen
- Ruta de La Vida 4 DemorasDokument44 SeitenRuta de La Vida 4 DemorasMario Fernando Villota Esparza0% (1)
- Anillo Waldeyer protección vías respiratoriasDokument3 SeitenAnillo Waldeyer protección vías respiratoriasBinxitOoh LokiitOoh PBNoch keine Bewertungen
- Guia de MicrobiologiaDokument80 SeitenGuia de MicrobiologiaHinO - Arturo0% (1)
- 1-12 Laberinto PosteriorDokument12 Seiten1-12 Laberinto Posteriorapi-3740897Noch keine Bewertungen
- Potenciales de Membrana y Potenciales de Accion PDFDokument53 SeitenPotenciales de Membrana y Potenciales de Accion PDFPerla LP100% (13)
- Datos de FiliaciónDokument13 SeitenDatos de FiliaciónYulyMeliNoch keine Bewertungen
- Fisiopatologia Respiratoria (Trastornos de Difusion)Dokument30 SeitenFisiopatologia Respiratoria (Trastornos de Difusion)Roger Alejandro MontoyaNoch keine Bewertungen
- Manual de Patologia GeneralDokument114 SeitenManual de Patologia GeneralCarolina PavezNoch keine Bewertungen
- PDF-Enfermedades de La Esclera y Conjuntiva-WordDokument20 SeitenPDF-Enfermedades de La Esclera y Conjuntiva-WordKabuki SamNoch keine Bewertungen
- Leucocitos PDFDokument25 SeitenLeucocitos PDFMarioNoch keine Bewertungen
- Manual de HistoDokument81 SeitenManual de HistoJulio JR Galaviz0% (2)
- Cuestionario de La Historia ClinicaDokument6 SeitenCuestionario de La Historia ClinicaLuisa EspinosaNoch keine Bewertungen
- Mediadores Quimicos de La Inflamacion y DolorDokument60 SeitenMediadores Quimicos de La Inflamacion y DolorDiana Quintero de Blasco100% (1)
- Carcinoma EmbrionarioDokument11 SeitenCarcinoma EmbrionarioEricka SumbaNoch keine Bewertungen
- Inflamación aguda y crónicaDokument6 SeitenInflamación aguda y crónicaRocio GallegosNoch keine Bewertungen
- Anatomia y Fisiologia OjoDokument38 SeitenAnatomia y Fisiologia OjoMariana TakataNoch keine Bewertungen
- Extraccion ADN (Sangre) IIDokument5 SeitenExtraccion ADN (Sangre) IISebastianNoch keine Bewertungen
- Práctica Dos Uso y Manejo Del Equipo de Laboratorio.Dokument10 SeitenPráctica Dos Uso y Manejo Del Equipo de Laboratorio.Cam PNoch keine Bewertungen
- ETIOPATOGENIADokument1 SeiteETIOPATOGENIALaura Mendoza AndradeNoch keine Bewertungen
- Patologia Inflamacion Mapa Conceptual. SaavedraDokument11 SeitenPatologia Inflamacion Mapa Conceptual. SaavedraJazmin Saavedra Mendoza86% (14)
- PDF 20230119 201952 0000Dokument21 SeitenPDF 20230119 201952 0000Karla JackeNoch keine Bewertungen
- Desequilibrios HidricosDokument10 SeitenDesequilibrios HidricosAndres AlmeidaNoch keine Bewertungen
- 1 PenicilinasDokument20 Seiten1 Penicilinasender-mendoza-6746Noch keine Bewertungen
- DNA polimerasas procariotas y eucariotasDokument6 SeitenDNA polimerasas procariotas y eucariotasRosario SandovalNoch keine Bewertungen
- Perfil de Lipidos 1Dokument28 SeitenPerfil de Lipidos 1GonzaloNoch keine Bewertungen
- Cascada de La CoagulaciónDokument26 SeitenCascada de La CoagulaciónDaniela León CastilloNoch keine Bewertungen
- Determinacion de FibrinogenoDokument7 SeitenDeterminacion de FibrinogenoAlejandra GualdronNoch keine Bewertungen
- Anatomía I: Disección del mediastino superior y posteriorDokument21 SeitenAnatomía I: Disección del mediastino superior y posteriorJhoselin Rojas ZentenoNoch keine Bewertungen
- Anticuerpos MonoclonalesDokument38 SeitenAnticuerpos MonoclonalesJose Luis Rivas CárdenasNoch keine Bewertungen
- Clasificacion de FracturasDokument56 SeitenClasificacion de FracturasCynthia CoutiñoNoch keine Bewertungen
- Exp. LinfopoyesisDokument9 SeitenExp. Linfopoyesisjuanbonifacio100% (4)
- Infecciones de Transmisión SexualDokument7 SeitenInfecciones de Transmisión SexualJavier García GuzmánNoch keine Bewertungen
- Cuestionario Sobre Trasplante de Organos y TejidosDokument3 SeitenCuestionario Sobre Trasplante de Organos y TejidosCecil OrtizNoch keine Bewertungen
- Cáncer de Hígado y de La EncrucijadaDokument5 SeitenCáncer de Hígado y de La EncrucijadaMonica Inspirit EXONFNTLNoch keine Bewertungen
- Español How To Use The VideofluorosDokument9 SeitenEspañol How To Use The VideofluorosDominique VegaNoch keine Bewertungen
- EBOOK Esenciales Del Hogar - 230209 - 224647Dokument14 SeitenEBOOK Esenciales Del Hogar - 230209 - 224647Fernando NavarroNoch keine Bewertungen
- Banco de Preguntas Semiologia 2. ECOE 2022Dokument13 SeitenBanco de Preguntas Semiologia 2. ECOE 2022Jhonatan Vasquez RojasNoch keine Bewertungen
- Producto Académico 2 PSICOFARMACOLOGIA 2Dokument3 SeitenProducto Académico 2 PSICOFARMACOLOGIA 2JOEL ANTONIO AVILA TORRESNoch keine Bewertungen
- Diapositiva Johanna6Dokument24 SeitenDiapositiva Johanna6Moiskel YisusNoch keine Bewertungen
- Qué Es La Discapacidad MúltipleDokument3 SeitenQué Es La Discapacidad MúltipleMARIA PAULA AYALA JIMENEZNoch keine Bewertungen
- HipertensionnnnDokument26 SeitenHipertensionnnnJose Ignacio Caceres ReyesNoch keine Bewertungen
- Estudio ComparativoDokument118 SeitenEstudio ComparativoFrancisco Ja ToNoch keine Bewertungen
- GPC OnicomicosisDokument34 SeitenGPC Onicomicosisdaniel martinez sandovalNoch keine Bewertungen
- Silabo Sistema Endocrino FinalDokument30 SeitenSilabo Sistema Endocrino FinalRogelioEucedaNoch keine Bewertungen
- Consentimiento InformadoDokument1 SeiteConsentimiento InformadoMiriam Orozco EscandónNoch keine Bewertungen
- Contusión cardiaca - lesiones y complicacionesDokument67 SeitenContusión cardiaca - lesiones y complicacionesRosalieNoch keine Bewertungen
- Monografias Aceites Esenciales XachDokument6 SeitenMonografias Aceites Esenciales XachZoe CarreónNoch keine Bewertungen
- Fiebre TifoideaDokument1 SeiteFiebre Tifoideamanuel nampulaNoch keine Bewertungen
- Sistema circulatorio: Corazón, arterias, venas y capilaresDokument9 SeitenSistema circulatorio: Corazón, arterias, venas y capilaresAna HuertasNoch keine Bewertungen
- Parafrasis Relacion Agente HospederoDokument6 SeitenParafrasis Relacion Agente HospederoAdiel CancheNoch keine Bewertungen
- Examen General de OrinaDokument5 SeitenExamen General de OrinaKAREN RODRIGUEZ BETANCOURTNoch keine Bewertungen
- HC Dr. CantelliDokument17 SeitenHC Dr. CantelliAnonymous atS3ugNoch keine Bewertungen
- Mapa Conceptual EmbarazoDokument3 SeitenMapa Conceptual EmbarazoUlises MarchenaNoch keine Bewertungen
- Adt-Ma-0001 Manual Programa Institucional de Farmacovigilancia Vigente Version 1Dokument42 SeitenAdt-Ma-0001 Manual Programa Institucional de Farmacovigilancia Vigente Version 1beatrNoch keine Bewertungen
- Componentes Sanguineos1Dokument17 SeitenComponentes Sanguineos1Carlos Osuna MuñozNoch keine Bewertungen
- Embarazo ProlongadoDokument4 SeitenEmbarazo ProlongadoJosé Luis GálvezNoch keine Bewertungen
- Manejo Anestesico Del Paciente TraumatizadoDokument12 SeitenManejo Anestesico Del Paciente Traumatizadoapi-3738847100% (3)
- Normas de Areas de EnfermeríaDokument7 SeitenNormas de Areas de EnfermeríaThommyNoch keine Bewertungen
- Analisis de Organigrama de Clinica PDFDokument38 SeitenAnalisis de Organigrama de Clinica PDFOliver López SNoch keine Bewertungen
- 05 La Oxigenoterapia en UrgenciasDokument5 Seiten05 La Oxigenoterapia en UrgenciasJohn PNoch keine Bewertungen