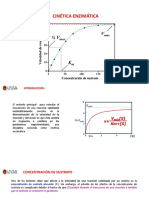
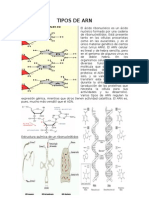
Das könnte Ihnen auch gefallen
- Presentacion 6 Cinetica EnzimaticaDokument46 SeitenPresentacion 6 Cinetica EnzimaticaE Clinton LMNoch keine Bewertungen
- Ejercicions No Mendel Agosto 2020Dokument2 SeitenEjercicions No Mendel Agosto 2020ellectrocompulsivoNoch keine Bewertungen
- Actividad EnzimaticaDokument8 SeitenActividad EnzimaticaLaura Franco SalazarNoch keine Bewertungen
- Reacciones MultisustratoDokument12 SeitenReacciones MultisustratoKarla Alejo GonzalezNoch keine Bewertungen
- Cuando El Bromuro de T-Butilo Se Calienta en Etanol A Reflujo Se Obtiene Una Mezzvla Formada Por: A) T-Butil Eter y Por 2-MetilpropenoDokument4 SeitenCuando El Bromuro de T-Butilo Se Calienta en Etanol A Reflujo Se Obtiene Una Mezzvla Formada Por: A) T-Butil Eter y Por 2-MetilpropenoFabricio JácomeNoch keine Bewertungen
- Tema 8 - Cinetica Enzimatica Farmacia PDFDokument12 SeitenTema 8 - Cinetica Enzimatica Farmacia PDFMariciell Larissa Gonzales TiconaNoch keine Bewertungen
- Bioenergética y MetabolismoDokument13 SeitenBioenergética y MetabolismoBerta González GómezNoch keine Bewertungen
- Marcadores MolecularesDokument10 SeitenMarcadores MolecularesJorge Isaac SanmiguelNoch keine Bewertungen
- ATP Sintasa-1Dokument15 SeitenATP Sintasa-1Omar BojorquezNoch keine Bewertungen
- Aplicaciones de Los Potenciales de Electrodo Estándar - g4Dokument49 SeitenAplicaciones de Los Potenciales de Electrodo Estándar - g4STEFHANY PAOLA MENDOZA HERNANDEZNoch keine Bewertungen
- Solucionario Taller de ProteinasDokument14 SeitenSolucionario Taller de ProteinasAndrés Felipe AcostaNoch keine Bewertungen
- Práctica 6 - Procesos Químicos Y Corriente Eléctrica - Aldo Esaú Xoxocotla SánchezDokument14 SeitenPráctica 6 - Procesos Químicos Y Corriente Eléctrica - Aldo Esaú Xoxocotla Sánchezxozo land100% (1)
- Reacciones de La Fase Luminosa de La FotosíntesisDokument3 SeitenReacciones de La Fase Luminosa de La FotosíntesisCarmen AvilaNoch keine Bewertungen
- Ejercicio 1 EnzimologiaDokument15 SeitenEjercicio 1 EnzimologiaAndrea ChavarroNoch keine Bewertungen
- 4efecto de La Actividad Enzimática de Inulinasas Usando Metales MG y Co Como Posibles Activador o Inhibidor Utilizando Kluyveromyces MarxianusDokument8 Seiten4efecto de La Actividad Enzimática de Inulinasas Usando Metales MG y Co Como Posibles Activador o Inhibidor Utilizando Kluyveromyces MarxianusEnrique GomezNoch keine Bewertungen
- Bioquimica Informe 3 UpsjbDokument24 SeitenBioquimica Informe 3 UpsjbJesus Flores Campos100% (2)
- Reglas de PaulingDokument9 SeitenReglas de PaulingMiguel Angel López NavarreteNoch keine Bewertungen
- Cromatografia para Purificación de ProteínasDokument55 SeitenCromatografia para Purificación de ProteínasAlejandro VieyraNoch keine Bewertungen
- Plegado de ProteinasDokument2 SeitenPlegado de Proteinastatiana toponNoch keine Bewertungen
- Replicación - KarpDokument14 SeitenReplicación - KarpCuarto Paralelo 1100% (1)
- Ruta MetabólicaDokument5 SeitenRuta MetabólicaKevin Guao BolañoNoch keine Bewertungen
- Poder Catalítico de Las EnzimasDokument16 SeitenPoder Catalítico de Las EnzimasAlex Chrome Dokuro-chan100% (1)
- Capitulos OrganicaDokument22 SeitenCapitulos OrganicaAlonsoAguasZuñigaNoch keine Bewertungen
- Ingenieria Metabolica de BacteriasDokument2 SeitenIngenieria Metabolica de BacteriasBrandon LopezNoch keine Bewertungen
- Informe 1 Aminoacido ArgininaDokument8 SeitenInforme 1 Aminoacido ArgininaAntonio MacNoch keine Bewertungen
- Estequiometria Microbiana 1Dokument5 SeitenEstequiometria Microbiana 1Richard Icaro EsparzaNoch keine Bewertungen
- Separacion de ProteinasDokument12 SeitenSeparacion de ProteinasAngie Juliana ParraNoch keine Bewertungen
- Gaden TraducidoDokument17 SeitenGaden TraducidoAdriana RamosNoch keine Bewertungen
- Biosintesis de AminoacidosDokument3 SeitenBiosintesis de AminoacidosKevinxdNoch keine Bewertungen
- Aplicaciones de La Clonación MolecularDokument3 SeitenAplicaciones de La Clonación MolecularAndrés Rodríguez MoleroNoch keine Bewertungen
- Efectos CooperativosDokument6 SeitenEfectos CooperativosJacs DenavNoch keine Bewertungen
- Conceptos Básicos de Cinética QuímicaDokument8 SeitenConceptos Básicos de Cinética QuímicaNicole Del Solar LagosNoch keine Bewertungen
- Minipreps, Tipos de Biotecnologia y Tipos de PromotoresDokument12 SeitenMinipreps, Tipos de Biotecnologia y Tipos de PromotoresManu RodriguezNoch keine Bewertungen
- Exposición de Ingeniería MetabólicaDokument29 SeitenExposición de Ingeniería MetabólicaEduardo Fabio Apari CossioNoch keine Bewertungen
- Examen ParcialDokument2 SeitenExamen ParcialElian Herrera Leon100% (1)
- Transcripcion de Eucariotas PDFDokument31 SeitenTranscripcion de Eucariotas PDFEduardo Mata GonzálezNoch keine Bewertungen
- Soluciones AmortiguadorasDokument6 SeitenSoluciones AmortiguadorasVivianaPiedraMontoyaNoch keine Bewertungen
- Efecto de Los Metales y Detergentes Sobre El CrecimientoDokument49 SeitenEfecto de Los Metales y Detergentes Sobre El CrecimientoEdgar Antonio Corro Islas0% (1)
- Reacciones EnzimaticasDokument9 SeitenReacciones EnzimaticasYesenia CuellarNoch keine Bewertungen
- PRÁCTICA NoDokument10 SeitenPRÁCTICA NoEmely Suhuchery Alfonso DiazNoch keine Bewertungen
- Hidrogenacion de Lípidos PDFDokument19 SeitenHidrogenacion de Lípidos PDFJose Bernardino RiveraNoch keine Bewertungen
- Fragmentacion PDFDokument22 SeitenFragmentacion PDFZai RSNoch keine Bewertungen
- Laboratorio Actividad EnzimaticaDokument7 SeitenLaboratorio Actividad EnzimaticaMaria Alejandra Diaz MejiaNoch keine Bewertungen
- Parametros BiológicosDokument26 SeitenParametros BiológicosCamila Sanabria0% (1)
- Estructura Acidos NucleicosDokument10 SeitenEstructura Acidos NucleicosTania TorresNoch keine Bewertungen
- 7 PKa de Un Indicador Acido BaseDokument8 Seiten7 PKa de Un Indicador Acido Basedaisuke_123Noch keine Bewertungen
- Tipos de ArnDokument5 SeitenTipos de ArnMiguel Angel Rodas Herrera100% (2)
- Termodinámica Ii Equilibrio Líquido Vapor en Un Sistema No IdealDokument8 SeitenTermodinámica Ii Equilibrio Líquido Vapor en Un Sistema No Idealyael vazquezNoch keine Bewertungen
- Balance EnergeticoDokument5 SeitenBalance EnergeticoGus RvNoch keine Bewertungen
- LIBbacarsDokument274 SeitenLIBbacarsMayte SalazarNoch keine Bewertungen
- Resumen de Regulacion A Nivel DNADokument11 SeitenResumen de Regulacion A Nivel DNALuis Antonio Perez AvitiaNoch keine Bewertungen
- 1 - Reacciones BisustratoDokument34 Seiten1 - Reacciones BisustratoKarla Dennise Guerra CruzNoch keine Bewertungen
- Propiedades Cinéticas de Las EnzimasDokument14 SeitenPropiedades Cinéticas de Las EnzimasMilagros Trujillo Mucha0% (1)
- Reacciones BisustratoDokument24 SeitenReacciones BisustratoMichelle HidroboNoch keine Bewertungen
- TEMA20Dokument46 SeitenTEMA20Mariana Linares QuevedoNoch keine Bewertungen
- Catalisis EnzimaticaDokument14 SeitenCatalisis EnzimaticaJessy Rivaz HerreraNoch keine Bewertungen
- Clase Tema 5a BiocatalisisDokument86 SeitenClase Tema 5a BiocatalisisArianaRiosNoch keine Bewertungen
- Cinética Enzimática - Parte 1 - Unidad 4Dokument62 SeitenCinética Enzimática - Parte 1 - Unidad 4Corazon ContentoNoch keine Bewertungen
- Tema2 EnzimasDokument5 SeitenTema2 EnzimasmorimbolaNoch keine Bewertungen
- Catalisis 2Dokument36 SeitenCatalisis 2ROSARIOMARTINEZ25Noch keine Bewertungen
- Calculo de Dietas Por Sistema de EquivalentesDokument37 SeitenCalculo de Dietas Por Sistema de EquivalentesRachell LoeraNoch keine Bewertungen
- Hematologia 3 - Serie Plaquetaria 2020Dokument35 SeitenHematologia 3 - Serie Plaquetaria 2020Angelo S VelNoch keine Bewertungen
- Tema 3 - LípidosDokument6 SeitenTema 3 - LípidosLara MaríaNoch keine Bewertungen
- Dokumen - Tips - Factores de Conversion VitaminasDokument2 SeitenDokumen - Tips - Factores de Conversion VitaminashermrzNoch keine Bewertungen
- Ruta de Las Pentosas FosfatoDokument10 SeitenRuta de Las Pentosas FosfatoAlexiiss LuiissNoch keine Bewertungen
- Diapositivas - Bioquimica - Acidos GrasosDokument18 SeitenDiapositivas - Bioquimica - Acidos GrasosMaryorisNoch keine Bewertungen
- Guia Trabajo N°6 Sem 2021-2Dokument3 SeitenGuia Trabajo N°6 Sem 2021-2Ayrton Lee Alfredo Aza�ero PerezNoch keine Bewertungen
- LIPASASDokument2 SeitenLIPASASliaNoch keine Bewertungen
- Purificacion de Una EnzimaDokument8 SeitenPurificacion de Una EnzimaAlex ReyesNoch keine Bewertungen
- Interacciones ADNDokument3 SeitenInteracciones ADNspike_5459Noch keine Bewertungen
- TRIGLICERIDODokument3 SeitenTRIGLICERIDOGabriel LeguaNoch keine Bewertungen
- Plasma Control Normal CoagulaciónDokument3 SeitenPlasma Control Normal CoagulaciónJose Andrés Parra LealNoch keine Bewertungen
- Propiedades de Los TriglicéridosDokument8 SeitenPropiedades de Los TriglicéridosCARLOS DIAZNoch keine Bewertungen
- Integración Metabólica en El Estado PostabsortivoDokument2 SeitenIntegración Metabólica en El Estado PostabsortivoangelikNoch keine Bewertungen
- Biología - George H. Fried - CarbohidratosDokument6 SeitenBiología - George H. Fried - CarbohidratosRODRÍGUEZ ALEJANDRONoch keine Bewertungen
- BIOQUIMICA DE HARPER Cap 51 Hemostacia y TrombosisDokument34 SeitenBIOQUIMICA DE HARPER Cap 51 Hemostacia y TrombosisJunior Quintero MoranNoch keine Bewertungen
- Membranas BiologicasDokument41 SeitenMembranas BiologicasJohanna Bustos NutricionistaNoch keine Bewertungen
- Ecuacion Michaelis y MentenDokument7 SeitenEcuacion Michaelis y MentenClase MicroNoch keine Bewertungen
- Todos Los Temas Curso 2020-2021Dokument204 SeitenTodos Los Temas Curso 2020-2021Sonia Hernández AbelairaNoch keine Bewertungen
- Lab 2. BioquimicaDokument27 SeitenLab 2. BioquimicaAngela Sofia Gonzalez HurtadoNoch keine Bewertungen
- Protein AsDokument5 SeitenProtein AsAngie OrdoñezNoch keine Bewertungen
- FosforilaciónDokument2 SeitenFosforilaciónCesar RenteriaNoch keine Bewertungen
- CITOEQUELETODokument3 SeitenCITOEQUELETOEliam G. SánchezNoch keine Bewertungen
- HORMONASDokument1 SeiteHORMONASRosalvina AquiseNoch keine Bewertungen
- 17 Citoesqueleto y MicrotúbulosDokument52 Seiten17 Citoesqueleto y MicrotúbulosJean VillanuevaNoch keine Bewertungen
- Taller EjerciciosDokument3 SeitenTaller EjerciciosValentina ArangoNoch keine Bewertungen
- Examen Biologia 3Dokument3 SeitenExamen Biologia 3marketingmundial100Noch keine Bewertungen
- Tarea 5. BM. Sanchez Mancillas. 2-06Dokument10 SeitenTarea 5. BM. Sanchez Mancillas. 2-06Beatriz Elena Sanchez MancillasNoch keine Bewertungen
- Lista Precios 2021 LubeDokument22 SeitenLista Precios 2021 LubeIsiPaz Scanner de VehiculosNoch keine Bewertungen
- Informe S9Dokument5 SeitenInforme S9Valeria DelpinoNoch keine Bewertungen
- Psicología oscura: Una guía esencial de persuasión, manipulación, engaño, control mental, negociación, conducta humana, PNL y guerra psicológicaVon EverandPsicología oscura: Una guía esencial de persuasión, manipulación, engaño, control mental, negociación, conducta humana, PNL y guerra psicológicaBewertung: 4.5 von 5 Sternen4.5/5 (766)
- Cómo hacer que te pasen cosas buenas: Entiende tu cerebro, gestiona tus emociones, mejora tu vidaVon EverandCómo hacer que te pasen cosas buenas: Entiende tu cerebro, gestiona tus emociones, mejora tu vidaBewertung: 5 von 5 Sternen5/5 (1875)
- Recupera tu mente, reconquista tu vidaVon EverandRecupera tu mente, reconquista tu vidaBewertung: 5 von 5 Sternen5/5 (8)
- Tus Zonas Erroneas: Guía Para Combatir las Causas de la InfelicidadVon EverandTus Zonas Erroneas: Guía Para Combatir las Causas de la InfelicidadBewertung: 4.5 von 5 Sternen4.5/5 (1833)
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceVon EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceBewertung: 5 von 5 Sternen5/5 (8)
- Yo Pude, ¡Tú Puedes!: Cómo tomar el control de tu bienestar emocional y convertirte en una persona imparable (edición revisada y expandida)Von EverandYo Pude, ¡Tú Puedes!: Cómo tomar el control de tu bienestar emocional y convertirte en una persona imparable (edición revisada y expandida)Bewertung: 5 von 5 Sternen5/5 (7)
- La violencia invisible: Identificar, entender y superar la violencia psicológica que sufrimos (y ejercemos) en nuestra vida cotidianaVon EverandLa violencia invisible: Identificar, entender y superar la violencia psicológica que sufrimos (y ejercemos) en nuestra vida cotidianaBewertung: 4 von 5 Sternen4/5 (2)
- El poder del optimismo: Herramientas para vivir de forma más positivaVon EverandEl poder del optimismo: Herramientas para vivir de forma más positivaBewertung: 5 von 5 Sternen5/5 (16)
- Resetea tu mente. Descubre de lo que eres capazVon EverandResetea tu mente. Descubre de lo que eres capazBewertung: 5 von 5 Sternen5/5 (196)
- La revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaVon EverandLa revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaBewertung: 5 von 5 Sternen5/5 (203)
- Cómo romper el ciclo de pensamientos negativosVon EverandCómo romper el ciclo de pensamientos negativosBewertung: 4.5 von 5 Sternen4.5/5 (274)
- Sistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Von EverandSistema nervioso y osteopatía: Nervios periféricos, meninges craneales y espinales, y sistema nervioso vegetativo (Color)Bewertung: 5 von 5 Sternen5/5 (9)
- Tu cerebro emocional: Saca partido de lo que sientes y transforma tu vidaVon EverandTu cerebro emocional: Saca partido de lo que sientes y transforma tu vidaBewertung: 5 von 5 Sternen5/5 (2)
- Resumen de Pensar rápido pensar despacio de Daniel KahnemanVon EverandResumen de Pensar rápido pensar despacio de Daniel KahnemanBewertung: 4.5 von 5 Sternen4.5/5 (64)
- Homo antecessor: El nacimiento de una especieVon EverandHomo antecessor: El nacimiento de una especieBewertung: 5 von 5 Sternen5/5 (1)
- Una mente en calma: Técnicas para manejar los pensamientos intrusivosVon EverandUna mente en calma: Técnicas para manejar los pensamientos intrusivosBewertung: 4.5 von 5 Sternen4.5/5 (142)
- Contra la ansiedad: Una guía completa para manejar emociones difícilesVon EverandContra la ansiedad: Una guía completa para manejar emociones difícilesBewertung: 5 von 5 Sternen5/5 (58)
- Batidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoVon EverandBatidos Verdes Depurativos y Antioxidantes: Aumenta tu Vitalidad con Smoothie Detox Durante 10 Días Para Adelgazar y Bajar de Peso: Aumenta tu vitalidad con smoothie detox durante 10 días para adelgazar y bajar de pesoBewertung: 5 von 5 Sternen5/5 (2)
- Escritura terapéutica. El poder curativo de la expresión escritaVon EverandEscritura terapéutica. El poder curativo de la expresión escritaBewertung: 5 von 5 Sternen5/5 (2)
- 50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaVon Everand50 técnicas de mindfulness para la ansiedad, la depresión, el estrés y el dolor: Mindfulness como terapiaBewertung: 4 von 5 Sternen4/5 (37)
- Influencia. La psicología de la persuasiónVon EverandInfluencia. La psicología de la persuasiónBewertung: 4.5 von 5 Sternen4.5/5 (14)
- Terapia cognitivo-conductual (TCC) y terapia dialéctico-conductual (TDC): Cómo la TCC, la TDC y la ACT pueden ayudarle a superar la ansiedad, la depresión, y los TOCSVon EverandTerapia cognitivo-conductual (TCC) y terapia dialéctico-conductual (TDC): Cómo la TCC, la TDC y la ACT pueden ayudarle a superar la ansiedad, la depresión, y los TOCSBewertung: 5 von 5 Sternen5/5 (1)