Das könnte Ihnen auch gefallen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
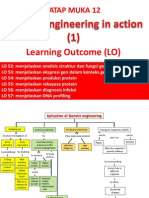
- TM 12 Genetic Engineering in Action 1 (Biologi Molekuler 2014)Dokument28 SeitenTM 12 Genetic Engineering in Action 1 (Biologi Molekuler 2014)himawarumNoch keine Bewertungen
- TM 03 Asam Nukleat (Biologi Molekuler 2014)Dokument23 SeitenTM 03 Asam Nukleat (Biologi Molekuler 2014)himawarumNoch keine Bewertungen
- TM 08 Polymerase Chain Reaction 2014Dokument17 SeitenTM 08 Polymerase Chain Reaction 2014himawarumNoch keine Bewertungen
- VirologiDokument28 SeitenVirologihimawarumNoch keine Bewertungen
- TM-08 Ethical Issues in Genetically Modified Organisms (GMOs) (Gasal 2014-2015)Dokument38 SeitenTM-08 Ethical Issues in Genetically Modified Organisms (GMOs) (Gasal 2014-2015)himawarumNoch keine Bewertungen
- Media Kultur Sel HewanDokument58 SeitenMedia Kultur Sel HewanhimawarumNoch keine Bewertungen
- 9a. Metabolisme Asam AminoDokument62 Seiten9a. Metabolisme Asam AminohimawarumNoch keine Bewertungen
- TM 12 Genetic Engineering in Action 1 (Biologi Molekuler 2014)Dokument28 SeitenTM 12 Genetic Engineering in Action 1 (Biologi Molekuler 2014)himawarumNoch keine Bewertungen
- TM 09 Selection, Screening, and Analysis of Recombinants 1 (2014)Dokument25 SeitenTM 09 Selection, Screening, and Analysis of Recombinants 1 (2014)ArdiantiNoch keine Bewertungen
- DIGESTIDokument110 SeitenDIGESTIhimawarumNoch keine Bewertungen
- DIGESTIDokument110 SeitenDIGESTIhimawarumNoch keine Bewertungen
- Nucleic AcidsDokument18 SeitenNucleic AcidshimawarumNoch keine Bewertungen
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (120)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Systematic Survey On Smart Home Safety and Security Systems Using The Arduino PlatformDokument24 SeitenSystematic Survey On Smart Home Safety and Security Systems Using The Arduino PlatformGeri MaeNoch keine Bewertungen
- The 7 Steps To Freedom - by David MacGregorDokument24 SeitenThe 7 Steps To Freedom - by David MacGregorTamás Dunavölgyi100% (1)
- Aarvee Denim & Export LTD.: CertificateDokument46 SeitenAarvee Denim & Export LTD.: CertificateVinita AgrawalNoch keine Bewertungen
- Lesson 2 Cultural Relativism - Part 1 (Reaction Paper)Dokument2 SeitenLesson 2 Cultural Relativism - Part 1 (Reaction Paper)Bai Zaida Abid100% (1)
- The Wall Dramaturgical PacketDokument24 SeitenThe Wall Dramaturgical PacketMarilo CostaNoch keine Bewertungen
- Dark Enemies by Effie CampbellDokument250 SeitenDark Enemies by Effie Campbellsheenuyadav06080Noch keine Bewertungen
- 332-Article Text-1279-1-10-20170327Dokument24 Seiten332-Article Text-1279-1-10-20170327Krisdayanti MendrofaNoch keine Bewertungen
- Spalding Application SampleDokument5 SeitenSpalding Application Sampleapi-66670156Noch keine Bewertungen
- Changing Views of Tattoos in JapanDokument14 SeitenChanging Views of Tattoos in JapanpanpapogNoch keine Bewertungen
- Interaksi Simbolik Dalam Komunikasi BudayaDokument18 SeitenInteraksi Simbolik Dalam Komunikasi BudayaPrazzNoch keine Bewertungen
- University of Mumbai: Bachelor of Management Studies (Finance) Semester VIDokument73 SeitenUniversity of Mumbai: Bachelor of Management Studies (Finance) Semester VIPranay ShettyNoch keine Bewertungen
- Psychological Science1Dokument23 SeitenPsychological Science1Jatin RaghavNoch keine Bewertungen
- Aron Nimzowitsch-My System & Chess Praxis-New in Chess (2016) PDFDokument1.029 SeitenAron Nimzowitsch-My System & Chess Praxis-New in Chess (2016) PDFMomin Fayzan100% (9)
- The Relationship Between Emotional Intelligence and Job Performance of Myanmar School TeachersDokument16 SeitenThe Relationship Between Emotional Intelligence and Job Performance of Myanmar School TeachersAmadeus Fernando M. PagenteNoch keine Bewertungen
- Thesis Project Management SoftwareDokument7 SeitenThesis Project Management Softwarehollyschulzgilbert100% (2)
- Law 431 Course Outline (Aug 22-23)Dokument3 SeitenLaw 431 Course Outline (Aug 22-23)Precious OmphithetseNoch keine Bewertungen
- Reflection Chapter 13 and 14Dokument2 SeitenReflection Chapter 13 and 14Vanessa Zevallos HernandezNoch keine Bewertungen
- Power Semiconductor DevicesDokument11 SeitenPower Semiconductor DevicesBhavesh Kaushal100% (1)
- Feminist Literary CriticismDokument11 SeitenFeminist Literary Criticismbela2011Noch keine Bewertungen
- Create and Open Input Dialog Box - MATLAB InputdlgDokument4 SeitenCreate and Open Input Dialog Box - MATLAB InputdlgwhytheritoNoch keine Bewertungen
- HRA Strategic PlanDokument40 SeitenHRA Strategic PlanjughomNoch keine Bewertungen
- Condition Monitoring 021711 v3Dokument40 SeitenCondition Monitoring 021711 v3Kevin_INoch keine Bewertungen
- Ass HeniDokument14 SeitenAss Henialexsabebe28Noch keine Bewertungen
- G8DDokument61 SeitenG8Dvivek1312100% (1)
- Impulse: Operation ManualDokument8 SeitenImpulse: Operation ManualRonggo SukmoNoch keine Bewertungen
- Experimental Nucleonics PDFDokument1 SeiteExperimental Nucleonics PDFEricNoch keine Bewertungen
- Disseminated Tuberculosis in An AIDS/HIV-Infected Patient: AbstractDokument3 SeitenDisseminated Tuberculosis in An AIDS/HIV-Infected Patient: AbstractAmelia Fitria DewiNoch keine Bewertungen
- WaveDokument5 SeitenWaveArijitSherlockMudiNoch keine Bewertungen
- Kerkythea Material Editor Guide 01Dokument36 SeitenKerkythea Material Editor Guide 01sapomasterNoch keine Bewertungen
- Catherine The Great: Catherine II, Empress of RussiaDokument7 SeitenCatherine The Great: Catherine II, Empress of RussiaLawrence James ParbaNoch keine Bewertungen