Das könnte Ihnen auch gefallen
- 7th Cytogenetics Lecture 20121 1Dokument37 Seiten7th Cytogenetics Lecture 20121 1RC SILVESTRENoch keine Bewertungen
- Eukaryotic Dna Replication 7 and 8Dokument81 SeitenEukaryotic Dna Replication 7 and 8Bintang PundhraNoch keine Bewertungen
- ICT in Education Enhancing Teaching and PDFDokument12 SeitenICT in Education Enhancing Teaching and PDFDebby Wura AbiodunNoch keine Bewertungen
- Health Through The Diet - An Ayurvedic Perspective - Pratibha Shah, BAMS, MD-AyuDokument40 SeitenHealth Through The Diet - An Ayurvedic Perspective - Pratibha Shah, BAMS, MD-AyuPriya Sharma SabaradNoch keine Bewertungen
- Almc Part - 2 Ayurvedic Lifestyle ManagementDokument101 SeitenAlmc Part - 2 Ayurvedic Lifestyle ManagementUdhayNoch keine Bewertungen
- Gene Cloning & Cloning Vectors: DR Ravi Kant Agrawal, MVSC, PHDDokument75 SeitenGene Cloning & Cloning Vectors: DR Ravi Kant Agrawal, MVSC, PHDvivekNoch keine Bewertungen
- Lac Operon: (A Negative Inducible Operon)Dokument32 SeitenLac Operon: (A Negative Inducible Operon)Alina RafiqueNoch keine Bewertungen
- Cell Cycle CampbellDokument67 SeitenCell Cycle CampbellSUNAYA GUSTI PUTUNoch keine Bewertungen
- Enzymes & Biological CatalysisDokument64 SeitenEnzymes & Biological Catalysisjoer13Noch keine Bewertungen
- Cell CycleDokument90 SeitenCell CycleSurneet HayerNoch keine Bewertungen
- PAPER 0703 Basics Concepts and Techniques in Molecular BiologyDokument70 SeitenPAPER 0703 Basics Concepts and Techniques in Molecular BiologyDiksha Gahlot100% (1)
- Regulation of Gene Expression in Prokaryotic and Eukaryotic CellsDokument13 SeitenRegulation of Gene Expression in Prokaryotic and Eukaryotic CellsMifta KhuljannahNoch keine Bewertungen
- 13 GeneRegulation Eukaryotes AMF S18 FullpageDokument37 Seiten13 GeneRegulation Eukaryotes AMF S18 FullpageSamuel LeNoch keine Bewertungen
- Erection Manual Hydro-1Dokument63 SeitenErection Manual Hydro-1rajfab100% (1)
- Genetics Chapter 16Dokument49 SeitenGenetics Chapter 16reed williamsNoch keine Bewertungen
- Protein and Amino AcidsDokument15 SeitenProtein and Amino AcidsArsalan khan100% (1)
- VectorsDokument56 SeitenVectorsShikha YashveerNoch keine Bewertungen
- Nucleus: Click To Edit Master Subtitle StyleDokument29 SeitenNucleus: Click To Edit Master Subtitle StyleAzifah ZakariaNoch keine Bewertungen
- Bacterial TransposonsDokument36 SeitenBacterial TransposonsShweta SinghNoch keine Bewertungen
- Cell Cycle RegulationDokument4 SeitenCell Cycle RegulationSomNoch keine Bewertungen
- Cell Cycle 4-09-09Dokument103 SeitenCell Cycle 4-09-09soundarya_madhiraNoch keine Bewertungen
- The Nuclear Pore ComplexDokument19 SeitenThe Nuclear Pore ComplexKunal SethNoch keine Bewertungen
- Porphyrins PDFDokument17 SeitenPorphyrins PDFPk JaatNoch keine Bewertungen
- Regulation of Gene Expression in Prokaryotes: © John Wiley & Sons, IncDokument65 SeitenRegulation of Gene Expression in Prokaryotes: © John Wiley & Sons, IncRoberto CastroNoch keine Bewertungen
- Body FluidsDokument38 SeitenBody FluidsGift AiyegbeniNoch keine Bewertungen
- CH17Dokument86 SeitenCH17y9bt429Noch keine Bewertungen
- Protein SynthesisDokument18 SeitenProtein Synthesisabisantiago6131100% (1)
- Medical Surgical Nursing-Ii 1Dokument51 SeitenMedical Surgical Nursing-Ii 1Shimmering MoonNoch keine Bewertungen
- HOMOLOGOUS AND NON HOMOLOGOUS RECOMBINATION by Preeti Saini 21mslsmm06Dokument7 SeitenHOMOLOGOUS AND NON HOMOLOGOUS RECOMBINATION by Preeti Saini 21mslsmm06Preeti SainiNoch keine Bewertungen
- Mach4 G and M Code Reference ManualDokument81 SeitenMach4 G and M Code Reference ManualMegi Setiawan SNoch keine Bewertungen
- Design of Torque ArmDokument16 SeitenDesign of Torque ArmRonak PanchalNoch keine Bewertungen
- Add Math Project Work 2013Dokument33 SeitenAdd Math Project Work 2013FrankieNgNoch keine Bewertungen
- Polymerase Chain Reaction: Salwa Hassan TeamaDokument56 SeitenPolymerase Chain Reaction: Salwa Hassan TeamaSamiksha SharmaNoch keine Bewertungen
- Change To ModbusTCP Slave R102Dokument8 SeitenChange To ModbusTCP Slave R102Евгений МалыченкоNoch keine Bewertungen
- Maintenance of 132 KV SwitchgearDokument34 SeitenMaintenance of 132 KV SwitchgearMuhammad Abdul Rauf100% (1)
- Answers PGR Week9 2016Dokument11 SeitenAnswers PGR Week9 2016kkk13whyNoch keine Bewertungen
- Answers PGR Week8Dokument3 SeitenAnswers PGR Week8kkk13whyNoch keine Bewertungen
- Blue-White SelectionDokument4 SeitenBlue-White SelectionBuse Nur UralNoch keine Bewertungen
- Lec 25Dokument17 SeitenLec 25john cenaNoch keine Bewertungen
- Gene Transfer MethodsDokument6 SeitenGene Transfer MethodsNawraj GhimireNoch keine Bewertungen
- Lambda Vectors and Their ReplicationDokument27 SeitenLambda Vectors and Their ReplicationSamridhi Singh100% (1)
- Genetic RecombinationDokument60 SeitenGenetic RecombinationJeevan BasnyatNoch keine Bewertungen
- Polymerase Chain Reaction (PCR)Dokument37 SeitenPolymerase Chain Reaction (PCR)Haleema SultanNoch keine Bewertungen
- BGY9003M Lecture 1 - Introduction 2017Dokument18 SeitenBGY9003M Lecture 1 - Introduction 2017jan0% (1)
- Chemical Method of Gene TransferDokument27 SeitenChemical Method of Gene TransferSreejesh P CNoch keine Bewertungen
- DR M.K.Bedi, Deptt. of Microbiology, DIBNSDokument35 SeitenDR M.K.Bedi, Deptt. of Microbiology, DIBNSRiya KashyapNoch keine Bewertungen
- Water and Life: Year 12 - Diploma BiologyDokument2 SeitenWater and Life: Year 12 - Diploma BiologyJohn OsborneNoch keine Bewertungen
- Intracellular Compartments and Protein Sorting: Reading: Alberts Et Al. 5th Edition 695-704, 721-727, 766-769, 779-783Dokument55 SeitenIntracellular Compartments and Protein Sorting: Reading: Alberts Et Al. 5th Edition 695-704, 721-727, 766-769, 779-783lemonpartymanNoch keine Bewertungen
- Internship Evaluation Form NewDokument3 SeitenInternship Evaluation Form Newsubhan sibghatNoch keine Bewertungen
- Staining Techniques in Biological StudiesDokument52 SeitenStaining Techniques in Biological StudiesRamyaAbhishekShenoyNoch keine Bewertungen
- Gene Mutation: Genomes 3Dokument72 SeitenGene Mutation: Genomes 3Abdus SubhanNoch keine Bewertungen
- Nucleus: Sudhanshu Shekhar, M.Tech BiotechDokument22 SeitenNucleus: Sudhanshu Shekhar, M.Tech BiotechSudhanshu ShekharNoch keine Bewertungen
- Eukaryote Regulation of Gene ExpressionDokument35 SeitenEukaryote Regulation of Gene ExpressiondewiulfaNoch keine Bewertungen
- Gene Transfer TechniquesDokument36 SeitenGene Transfer TechniquesDrMumtaz F MusaliarNoch keine Bewertungen
- Bacteriology Lab 2002Dokument89 SeitenBacteriology Lab 2002Romanda GreeneNoch keine Bewertungen
- TransposonsDokument6 SeitenTransposonsnikhilsathwikNoch keine Bewertungen
- Animal Tissue CultureDokument23 SeitenAnimal Tissue CultureHui Jun Hoe80% (5)
- Blue WhiteDokument2 SeitenBlue WhiteShyamala RamalingamNoch keine Bewertungen
- Atomic Force MicrosDokument26 SeitenAtomic Force MicrosGnana Sri Durga ParuchuriNoch keine Bewertungen
- Mechanism of Protein Targeting Into Endoplasmic ReticulumDokument24 SeitenMechanism of Protein Targeting Into Endoplasmic ReticulumRebati Raman PandaNoch keine Bewertungen
- Translation in ProkaryotesDokument25 SeitenTranslation in ProkaryotesNaomi Lamtiur0% (1)
- Cell Cycle Checkpoints PPT by EasybiologyclassDokument22 SeitenCell Cycle Checkpoints PPT by EasybiologyclassMaryNoch keine Bewertungen
- Confocal Microscopy: DR R.JayapradaDokument23 SeitenConfocal Microscopy: DR R.JayapradaprinceamitNoch keine Bewertungen
- Applications of Genetic Engineering in AgricultureDokument10 SeitenApplications of Genetic Engineering in AgricultureDeepika KVNoch keine Bewertungen
- Chapter 3b Worksheet OpenstaxDokument10 SeitenChapter 3b Worksheet Openstaxapi-255334265Noch keine Bewertungen
- Group Two'S Seminar Work: Topic: Enzyme Regulation Allosteric Regulation and Models OutlineDokument13 SeitenGroup Two'S Seminar Work: Topic: Enzyme Regulation Allosteric Regulation and Models OutlineOluwasegun ModupeNoch keine Bewertungen
- Afik FixDokument1 SeiteAfik FixWahyu KurniawatiNoch keine Bewertungen
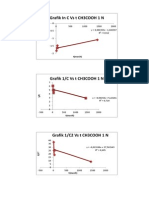
- Grafik in C Vs T CH3COOH 1 NDokument2 SeitenGrafik in C Vs T CH3COOH 1 NWahyu KurniawatiNoch keine Bewertungen
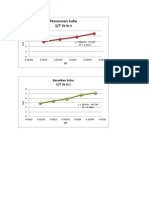
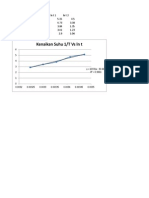
- Kenaikan Suhu 1/T Vs LN TDokument2 SeitenKenaikan Suhu 1/T Vs LN TWahyu KurniawatiNoch keine Bewertungen
- Are Humans Responsible For Global Warming ?Dokument4 SeitenAre Humans Responsible For Global Warming ?Wahyu KurniawatiNoch keine Bewertungen
- Important Notes of 10th Class Math Exercise 3.2Dokument12 SeitenImportant Notes of 10th Class Math Exercise 3.2Tayyabah ShahNoch keine Bewertungen
- Tutorial On SchedulingDokument14 SeitenTutorial On Schedulingrktiwary256034Noch keine Bewertungen
- Lab 1Dokument4 SeitenLab 1zakariaNoch keine Bewertungen
- Ultima X Series Instruction Manual - enDokument141 SeitenUltima X Series Instruction Manual - enStefano EsmNoch keine Bewertungen
- Handout E.15 - Examples On Transient Response of First and Second Order Systems, System Damping and Natural FrequencyDokument14 SeitenHandout E.15 - Examples On Transient Response of First and Second Order Systems, System Damping and Natural FrequencyRishikesh BhavsarNoch keine Bewertungen
- Sci Problem SolutionsDokument10 SeitenSci Problem SolutionsVS PUBLIC SCHOOL BangaloreNoch keine Bewertungen
- Dot 57406 DS1Dokument172 SeitenDot 57406 DS1Carlos FernandezNoch keine Bewertungen
- Rapid Prototyping PPT SeminarDokument32 SeitenRapid Prototyping PPT SeminarShantha Kumar G C0% (1)
- New Microsoft Office Word DocumentDokument20 SeitenNew Microsoft Office Word DocumentVinay KumarNoch keine Bewertungen
- Chapter 6 - Ion ChannelsDokument12 SeitenChapter 6 - Ion ChannelsntghshrNoch keine Bewertungen
- Python (Dictionary)Dokument54 SeitenPython (Dictionary)shahrukhkr.gptNoch keine Bewertungen
- CHEM101 051 Old-Exam Second-Major Master-KeyDokument10 SeitenCHEM101 051 Old-Exam Second-Major Master-KeyalwafiNoch keine Bewertungen
- BPCL Training ReportDokument34 SeitenBPCL Training ReportVishalVaishNoch keine Bewertungen
- Leadership Quality ManagementDokument8 SeitenLeadership Quality Managementselinasimpson361Noch keine Bewertungen
- Addressable Loop Isolator Module AI 520Dokument2 SeitenAddressable Loop Isolator Module AI 520ராஜா கிருஷ்ணன்Noch keine Bewertungen
- Engineering MeasurementsDokument77 SeitenEngineering MeasurementsFemi Prince0% (1)
- Structure of Atom - Class 11thDokument38 SeitenStructure of Atom - Class 11thAdil KhanNoch keine Bewertungen
- In The Next Three Chapters, We Will Examine Different Aspects of Capital Market Theory, IncludingDokument62 SeitenIn The Next Three Chapters, We Will Examine Different Aspects of Capital Market Theory, IncludingRahmat M JayaatmadjaNoch keine Bewertungen
- Rr212203 Transducers and Instrumentation ComponentsDokument8 SeitenRr212203 Transducers and Instrumentation ComponentsSrinivasa Rao GNoch keine Bewertungen
- Learning Objectives: Lecture 3: Moving Averages and Exponential SmoothingDokument6 SeitenLearning Objectives: Lecture 3: Moving Averages and Exponential SmoothingcuriousdumboNoch keine Bewertungen
- A Handheld Gun Detection Using Faster R-CNN Deep LearningDokument5 SeitenA Handheld Gun Detection Using Faster R-CNN Deep LearningJanderson LiraNoch keine Bewertungen
- 6PC Arrears CalculatorDokument6 Seiten6PC Arrears Calculatoranon-530704Noch keine Bewertungen
- Sen2022 Software Engineering Analysis and DesignDokument84 SeitenSen2022 Software Engineering Analysis and DesignElif KaraatlıNoch keine Bewertungen