Das könnte Ihnen auch gefallen
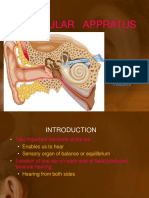
- Vestibular Appratus: BY: Harshita YadavDokument49 SeitenVestibular Appratus: BY: Harshita YadavNitin SharmaNoch keine Bewertungen
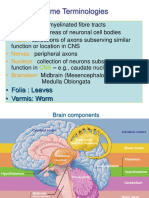
- Cerebellum and Brain Stem: DR Asim Shrestha SRCC Ped Neuro Fellow MumbaiDokument71 SeitenCerebellum and Brain Stem: DR Asim Shrestha SRCC Ped Neuro Fellow MumbaiAsim ShresthaNoch keine Bewertungen
- 3-Ascending Tracts of Spinal CordDokument28 Seiten3-Ascending Tracts of Spinal Cordmuhammad altaf100% (2)
- Cerebellum: Principally Because Electrical Excitation of The CerebellumDokument42 SeitenCerebellum: Principally Because Electrical Excitation of The Cerebellumdr_mksinhaNoch keine Bewertungen
- Physiology of Hearing & BalanceDokument41 SeitenPhysiology of Hearing & BalanceSudhir BayyaNoch keine Bewertungen
- Understanding A Dizzy AdultDokument39 SeitenUnderstanding A Dizzy AdultR Srinivasa RaghavanNoch keine Bewertungen
- Physiology of Hearing and Equilibrium LectureDokument52 SeitenPhysiology of Hearing and Equilibrium LectureHomeground entertainment100% (1)
- Cholesteatoma FinalDokument56 SeitenCholesteatoma FinalDr.Bharathi M.BNoch keine Bewertungen
- Head & Neck - Deep Cervical Fascia &cervical PlexusDokument9 SeitenHead & Neck - Deep Cervical Fascia &cervical PlexusKomang Tri Adi SuparwatiNoch keine Bewertungen
- Cerebellum, Thalamus, and HypothalamusDokument40 SeitenCerebellum, Thalamus, and Hypothalamusasep100% (1)
- Vertebral Arteries, and Their Divisions. Arteries Fuse To Form The Basilar ArteryDokument6 SeitenVertebral Arteries, and Their Divisions. Arteries Fuse To Form The Basilar Arterymurali_bharadwazNoch keine Bewertungen
- PHYSIOLOGY OF BALANCE: A MULTISENSORY INTEGRATION PROCESSDokument18 SeitenPHYSIOLOGY OF BALANCE: A MULTISENSORY INTEGRATION PROCESSKunal ShahNoch keine Bewertungen
- Erb's PointDokument14 SeitenErb's PointRohit SharmaNoch keine Bewertungen
- Nerve Conduction Velocity: Karunakaran K MPT (Neuro) ., PGDMT Associate Professor, SvcoptDokument30 SeitenNerve Conduction Velocity: Karunakaran K MPT (Neuro) ., PGDMT Associate Professor, SvcoptSineka 19170239Noch keine Bewertungen
- ANATOMY AND PHYSIOLOGY OF THE VESTIBULAR ORGAN AND NEURAL PATHWAYSDokument56 SeitenANATOMY AND PHYSIOLOGY OF THE VESTIBULAR ORGAN AND NEURAL PATHWAYSVineet Chadha0% (1)
- Neurological ExaminationDokument240 SeitenNeurological Examinationramadan0% (1)
- Cranial Neuropathies Guide: Electrodiagnosis and ManagementDokument74 SeitenCranial Neuropathies Guide: Electrodiagnosis and ManagementKevin Woodard100% (1)
- A0618 Hearing and EquilibriumDokument19 SeitenA0618 Hearing and EquilibriumSeptiani HasibuanNoch keine Bewertungen
- k4 - Disorders of Cranial NervesDokument67 Seitenk4 - Disorders of Cranial Nerveswlmhfp100% (1)
- Causes and Management of Syncope in DentistryDokument27 SeitenCauses and Management of Syncope in DentistrySelvarathi KandhaswamyNoch keine Bewertungen
- Anatomy of Vestibular SystemDokument25 SeitenAnatomy of Vestibular SystemFranciscus BuwanaNoch keine Bewertungen
- Anatomy of the Hypopharynx and LarynxDokument39 SeitenAnatomy of the Hypopharynx and LarynxArjun NambiarNoch keine Bewertungen
- Motor System: DR - Ahmed Gaber Ass. Prof of Neurology Ain Shams UniversityDokument63 SeitenMotor System: DR - Ahmed Gaber Ass. Prof of Neurology Ain Shams UniversityKhaled Ossama100% (1)
- Facial Nerve Palsy: Dr. Saud AlromaihDokument74 SeitenFacial Nerve Palsy: Dr. Saud AlromaihChandra ManapaNoch keine Bewertungen
- BPPVDokument10 SeitenBPPVRizki MaulanaNoch keine Bewertungen
- Hold the foot firmly in dorsiflexion to relax the gastrocnemiusmuscle and expose the Achilles tendon fullyDokument13 SeitenHold the foot firmly in dorsiflexion to relax the gastrocnemiusmuscle and expose the Achilles tendon fullyNurul Huda M ShahrinNoch keine Bewertungen
- Rare spinal cord disease syringomyelia definition causes symptoms treatmentDokument8 SeitenRare spinal cord disease syringomyelia definition causes symptoms treatmentNurfadiyah YasinNoch keine Bewertungen
- Sphenoid BoneDokument76 SeitenSphenoid BoneLALITH SAI KNoch keine Bewertungen
- Anatomy & Physiology of Ear (Prof. Fatthi Abdel-Baki)Dokument20 SeitenAnatomy & Physiology of Ear (Prof. Fatthi Abdel-Baki)Dr-Firas Nayf Al-ThawabiaNoch keine Bewertungen
- Vestibular System Anatomy and FunctionDokument7 SeitenVestibular System Anatomy and FunctionKevinYonathanNoch keine Bewertungen
- Vestibular DisordersDokument8 SeitenVestibular DisordersHilwy Al-haninNoch keine Bewertungen
- 21-Spinal Cord TractsDokument23 Seiten21-Spinal Cord TractsALFAHRUL CAHYADINoch keine Bewertungen
- Dura Mater & Cranial SinusDokument30 SeitenDura Mater & Cranial SinusSadiq Wadood SiddiquiNoch keine Bewertungen
- Cerebrum Sulci and Gyri and Circle of WillisDokument53 SeitenCerebrum Sulci and Gyri and Circle of Willisgandhiayu100% (1)
- Assr, Vemp, VNG, OaeDokument8 SeitenAssr, Vemp, VNG, OaeSuprit Sn100% (1)
- Malignant Otitis Externa 20070821Dokument0 SeitenMalignant Otitis Externa 20070821Suci PramadianiNoch keine Bewertungen
- Anatomy of Labyrinth by DR Inam Ur RehmanDokument29 SeitenAnatomy of Labyrinth by DR Inam Ur RehmanasssadulllahNoch keine Bewertungen
- NeurologyDokument149 SeitenNeurologyAimee100% (1)
- STRETCH REFLEX ANALYSISDokument31 SeitenSTRETCH REFLEX ANALYSISTots Enriquez Navarro100% (1)
- Anatomy of Cerebral Blood Supply & Cerebral PhysiologyDokument32 SeitenAnatomy of Cerebral Blood Supply & Cerebral Physiologykiran kizhakkiniNoch keine Bewertungen
- Head-Shaking Nystagmus (04.04.2005)Dokument11 SeitenHead-Shaking Nystagmus (04.04.2005)r.boniver524Noch keine Bewertungen
- Nerves of Upper LimbDokument33 SeitenNerves of Upper LimbAamir ShahzadNoch keine Bewertungen
- Cerebellar Ataxia Pathophysiology and RehabilitationDokument23 SeitenCerebellar Ataxia Pathophysiology and RehabilitationMikail AtiyehNoch keine Bewertungen
- Spinal Cord LesionsDokument110 SeitenSpinal Cord Lesionsgtaha80100% (1)
- Review of Anatomy of The EarDokument16 SeitenReview of Anatomy of The EarSahrish IqbalNoch keine Bewertungen
- Anatomy of Spinal CordDokument47 SeitenAnatomy of Spinal CordmayseeNoch keine Bewertungen
- Lung SurgeriesDokument43 SeitenLung SurgeriesSereinNoch keine Bewertungen
- Auditory Brainstem ResponseDokument18 SeitenAuditory Brainstem ResponseFree dataNoch keine Bewertungen
- Internal CapsuleDokument25 SeitenInternal CapsuleMadan KumarNoch keine Bewertungen
- The Ventricular SystemDokument6 SeitenThe Ventricular SystemM Arsalan TariqNoch keine Bewertungen
- 2022 OlfactionDokument46 Seiten2022 OlfactionRam LakshmanNoch keine Bewertungen
- Anat 6.6 Cerebellum - DeriadaDokument11 SeitenAnat 6.6 Cerebellum - Deriadalovelots1234Noch keine Bewertungen
- Neural Mechanisms of Salivary Gland SecretionDokument247 SeitenNeural Mechanisms of Salivary Gland SecretionMassimo Baudo100% (1)
- Lower Motor Neuron (LMN) Facial Palsy: Department of ENTDokument32 SeitenLower Motor Neuron (LMN) Facial Palsy: Department of ENTalfaz lakhani100% (1)
- ASIA ScoreDokument14 SeitenASIA Scorezloncar3Noch keine Bewertungen
- Essential Clinically Applied Anatomy of the Peripheral Nervous System in the Head and NeckVon EverandEssential Clinically Applied Anatomy of the Peripheral Nervous System in the Head and NeckNoch keine Bewertungen
- Consultant in Audiology: Passbooks Study GuideVon EverandConsultant in Audiology: Passbooks Study GuideNoch keine Bewertungen
- Multiple Choice Questions in Ophthalmic and NeuroanatomyVon EverandMultiple Choice Questions in Ophthalmic and NeuroanatomyNoch keine Bewertungen
- Declarations of High Self Esteem PDFDokument2 SeitenDeclarations of High Self Esteem PDFsharkss521Noch keine Bewertungen
- The Discrimination ModelDokument16 SeitenThe Discrimination ModelSiti MuslihaNoch keine Bewertungen
- Exam Per DevDokument2 SeitenExam Per DevXexArias100% (2)
- Biophysics Assignment2 2k19ep042 Hrithik KalkalDokument4 SeitenBiophysics Assignment2 2k19ep042 Hrithik KalkalHarsh Raj EP-037Noch keine Bewertungen
- The Relationship Between Perceived Inter-Parental Conflict and Academic Adjustment in First Year Students' Hamadan, IranDokument5 SeitenThe Relationship Between Perceived Inter-Parental Conflict and Academic Adjustment in First Year Students' Hamadan, IranIsabel AntunesNoch keine Bewertungen
- Sample Training Course OutlineDokument3 SeitenSample Training Course OutlineKRISHNANNoch keine Bewertungen
- BelieveDokument2 SeitenBelieveArmy729Noch keine Bewertungen
- Assessing Stress Management of Criminology StudentsDokument20 SeitenAssessing Stress Management of Criminology StudentsDick Jefferson Ocampo PatingNoch keine Bewertungen
- The Dangers of All-NightersDokument2 SeitenThe Dangers of All-Nightersadhiimull gantengNoch keine Bewertungen
- Human Social Temperaments: Yin, Yang, and HarmonyDokument32 SeitenHuman Social Temperaments: Yin, Yang, and Harmonydao einsnewt100% (2)
- Sample Business MemoDokument1 SeiteSample Business MemoLynlie PeraltaNoch keine Bewertungen
- Sunrise Sunrise: zb1) Z b2)Dokument26 SeitenSunrise Sunrise: zb1) Z b2)jn875849Noch keine Bewertungen
- Dreams and Dreaming (Stanford Encyclopedia of Philosophy)Dokument28 SeitenDreams and Dreaming (Stanford Encyclopedia of Philosophy)paul tNoch keine Bewertungen
- Panic Disorder PhobiasDokument9 SeitenPanic Disorder Phobiasapi-3797941Noch keine Bewertungen
- Spatial Cognition Libro 1998Dokument383 SeitenSpatial Cognition Libro 1998JuanpeGarbayoNoch keine Bewertungen
- Siamese Network: Shusen WangDokument51 SeitenSiamese Network: Shusen WangMInh ThanhNoch keine Bewertungen
- Neurospa - Manual 2016 - 220829 - 202650Dokument8 SeitenNeurospa - Manual 2016 - 220829 - 202650Liza HaeberleNoch keine Bewertungen
- INTERPERSONAL BEHAVIOUR MANAGEMENTDokument27 SeitenINTERPERSONAL BEHAVIOUR MANAGEMENTMohd FadhlyNoch keine Bewertungen
- What Is Geographical Enquiry?Dokument11 SeitenWhat Is Geographical Enquiry?geographyrocks100% (1)
- Neural Devices and Neurosecurity Final Paper2Dokument7 SeitenNeural Devices and Neurosecurity Final Paper2api-570817284Noch keine Bewertungen
- L A 12 and S Q 12Dokument7 SeitenL A 12 and S Q 12api-374915232Noch keine Bewertungen
- 7.cimt & MRPDokument41 Seiten7.cimt & MRPOsama Malik0% (1)
- Call of Cthulhu 7e - Core RulebookDokument4 SeitenCall of Cthulhu 7e - Core RulebookClaudio Andrés Artigas0% (2)
- Unit 4 Written Assiignment RDokument4 SeitenUnit 4 Written Assiignment RJackson LtorishaNoch keine Bewertungen
- GROUP REPORT PATOFISIOLOGI EnglishDokument18 SeitenGROUP REPORT PATOFISIOLOGI EnglishIndo PutraNoch keine Bewertungen
- Posttraumatic Growth in Youth Survivors of A DisasterDokument9 SeitenPosttraumatic Growth in Youth Survivors of A DisasterLakshmi ShankarNoch keine Bewertungen
- Chapter - Ii: Review of LiteratureDokument4 SeitenChapter - Ii: Review of LiteratureSalman RazaNoch keine Bewertungen
- Emotional Intelligence - When Smart Is DumbDokument2 SeitenEmotional Intelligence - When Smart Is DumbArchana ChhatparNoch keine Bewertungen
- Bruce Ecker, Laurel Hulley - Coherence Therapy - Practice Manual & Training Guide-Coherence Psychology Institute LLC (2017)Dokument93 SeitenBruce Ecker, Laurel Hulley - Coherence Therapy - Practice Manual & Training Guide-Coherence Psychology Institute LLC (2017)John Smith67% (6)
- DLL Week 4 Q3 NMSDokument11 SeitenDLL Week 4 Q3 NMSCrystal NideaNoch keine Bewertungen