Das könnte Ihnen auch gefallen
- Plantilla Pre-Venta - ValidaciónDokument2 SeitenPlantilla Pre-Venta - ValidaciónKarolLeylaNoch keine Bewertungen
- Programacion Enero 2022 ComplementariasDokument3 SeitenProgramacion Enero 2022 ComplementariasKarolLeylaNoch keine Bewertungen
- Newsletter 130 Supplement SpaDokument12 SeitenNewsletter 130 Supplement SpaKarolLeylaNoch keine Bewertungen
- Taller de Protección Respiratoria LaboralDokument28 SeitenTaller de Protección Respiratoria LaboralKarolLeylaNoch keine Bewertungen
- 40-Texto Del Artículo-114-1-10-20210607Dokument21 Seiten40-Texto Del Artículo-114-1-10-20210607KarolLeylaNoch keine Bewertungen
- Proyeccion Social MedicinaDokument13 SeitenProyeccion Social MedicinaKarolLeylaNoch keine Bewertungen
- CartaDokument2 SeitenCartaKarolLeylaNoch keine Bewertungen
- INFLUENZANUEVOIMPORTANTE2010Dokument2 SeitenINFLUENZANUEVOIMPORTANTE2010KarolLeylaNoch keine Bewertungen
- Newsletter 130 Supplement SpaDokument12 SeitenNewsletter 130 Supplement SpaKarolLeylaNoch keine Bewertungen
- ANTIBIÓTICOS PEDIATRÍA: DROGAS Y MECANISMOS DE ACCIÓNDokument21 SeitenANTIBIÓTICOS PEDIATRÍA: DROGAS Y MECANISMOS DE ACCIÓNKarolLeylaNoch keine Bewertungen
- Fcs-fr-30 Declaracion Jurada - Evaluacion de Proyecto de InvestigacionDokument2 SeitenFcs-fr-30 Declaracion Jurada - Evaluacion de Proyecto de InvestigacionKarolLeylaNoch keine Bewertungen
- Signos Vitales:: Hora de La Atención Profesional Asistencial Especialidad InterconsultaDokument2 SeitenSignos Vitales:: Hora de La Atención Profesional Asistencial Especialidad InterconsultaKarolLeylaNoch keine Bewertungen
- j5 Examen Seminario IIDokument30 Seitenj5 Examen Seminario IIKarolLeylaNoch keine Bewertungen
- Automed IntDokument74 SeitenAutomed IntKarolLeylaNoch keine Bewertungen
- Caso 1 VLRDokument10 SeitenCaso 1 VLRKarolLeylaNoch keine Bewertungen
- Automed IntDokument74 SeitenAutomed IntKarolLeylaNoch keine Bewertungen
- Manejo de Caso de IntoxicaciónDokument8 SeitenManejo de Caso de IntoxicaciónKarolLeylaNoch keine Bewertungen
- Banco de Preguntas ENARM (600 Preguntas - Contestadas) PDFDokument217 SeitenBanco de Preguntas ENARM (600 Preguntas - Contestadas) PDFHugo Emilio Naupa Medina50% (2)
- Manejo de Caso de Sintomatico RespiratorioDokument41 SeitenManejo de Caso de Sintomatico RespiratorioKarolLeylaNoch keine Bewertungen
- Diferenciacion SexualDokument38 SeitenDiferenciacion SexualyolcharnnguenNoch keine Bewertungen
- Signos Vitales:: Hora de La Atención Profesional Asistencial Especialidad InterconsultaDokument2 SeitenSignos Vitales:: Hora de La Atención Profesional Asistencial Especialidad InterconsultaKarolLeylaNoch keine Bewertungen
- Signos Vitales:: Hora de La Atención Profesional Asistencial Especialidad InterconsultaDokument2 SeitenSignos Vitales:: Hora de La Atención Profesional Asistencial Especialidad InterconsultaKarolLeylaNoch keine Bewertungen
- PROYECTO DE INVESTIGACION ModeloDokument10 SeitenPROYECTO DE INVESTIGACION ModeloKarolLeylaNoch keine Bewertungen
- Manejo de Caso de Sintomatico RespiratorioDokument41 SeitenManejo de Caso de Sintomatico RespiratorioKarolLeylaNoch keine Bewertungen
- HC ACV HemorragicoDokument14 SeitenHC ACV HemorragicoKarolLeylaNoch keine Bewertungen
- Manejo de Caso de Dolor ToracicoDokument12 SeitenManejo de Caso de Dolor ToracicoKarolLeylaNoch keine Bewertungen
- Manejo de Caso de Sindrome Doloroso AbdominalDokument9 SeitenManejo de Caso de Sindrome Doloroso AbdominalKarolLeylaNoch keine Bewertungen
- Carta de CotizacionDokument1 SeiteCarta de CotizacionDilma GomezNoch keine Bewertungen
- Manejo de Caso de IntoxicaciónDokument8 SeitenManejo de Caso de IntoxicaciónKarolLeylaNoch keine Bewertungen
- Modelo+de+solicitud+de+cotización+no +1+ (Bienes)Dokument1 SeiteModelo+de+solicitud+de+cotización+no +1+ (Bienes)MarioJoseTelcomNoch keine Bewertungen
- Hipercolesterolemia Familiar: Causas, Síntomas y Tratamiento con EstatinasDokument10 SeitenHipercolesterolemia Familiar: Causas, Síntomas y Tratamiento con EstatinasFelipe Javier Melis ArcosNoch keine Bewertungen
- Informe 2 Practica de Laboratorio PDFDokument5 SeitenInforme 2 Practica de Laboratorio PDFLuis CanoNoch keine Bewertungen
- Manual de Intervención en Crisis en Situaciones de Desastre PDFDokument38 SeitenManual de Intervención en Crisis en Situaciones de Desastre PDFDr. José Antonio Ayerdis ZamoraNoch keine Bewertungen
- Composición Del HuesoDokument12 SeitenComposición Del HuesoTessy Cueva QuevedoNoch keine Bewertungen
- Como Iniciar Con Un AcuarioDokument5 SeitenComo Iniciar Con Un AcuarioVictor Olvera RochaNoch keine Bewertungen
- Cinetica LipasasDokument4 SeitenCinetica LipasasGabriel LVNoch keine Bewertungen
- Trabajo de Investigacion de La ShainaDokument7 SeitenTrabajo de Investigacion de La ShainaBetzi EmperatrizNoch keine Bewertungen
- 401 Alteraciones EquilibrioDokument7 Seiten401 Alteraciones EquilibrioLisset AdasmeNoch keine Bewertungen
- Principales variables en mecánica respiratoriaDokument33 SeitenPrincipales variables en mecánica respiratoriaJorge PérezNoch keine Bewertungen
- Focusing DibujadoDokument11 SeitenFocusing DibujadoGonzalo Abarzúa100% (1)
- Eter NaftilmetilicoDokument6 SeitenEter Naftilmetilicodavid torresNoch keine Bewertungen
- Encefalopatia Espongiforme BovinaDokument45 SeitenEncefalopatia Espongiforme BovinaHM BelphegorNoch keine Bewertungen
- El Timo y Sus GeneralidadesDokument5 SeitenEl Timo y Sus GeneralidadesGiane VacaNoch keine Bewertungen
- Libro para colorear aves del Parque de las AvesDokument80 SeitenLibro para colorear aves del Parque de las AvesPepita Pérez100% (1)
- Museo de Medicina EnsayoDokument4 SeitenMuseo de Medicina EnsayoMichelle MarinNoch keine Bewertungen
- Guia 1 - Laboratorio Microbiologia.Dokument8 SeitenGuia 1 - Laboratorio Microbiologia.KiuzNoch keine Bewertungen
- Tesis Memoria en Adultos.Dokument201 SeitenTesis Memoria en Adultos.Alfonso Yunge-MorinNoch keine Bewertungen
- 1 Concepto Materia Configuracion Propiedades PeriodicasDokument58 Seiten1 Concepto Materia Configuracion Propiedades PeriodicasMabel Elisa Rodriguez MendezNoch keine Bewertungen
- Banco de Preguntas - Esfm - 2018Dokument93 SeitenBanco de Preguntas - Esfm - 2018Franz Ever Quispecahuana Limachi80% (111)
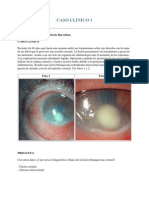
- Caso Clinico OftalmoDokument20 SeitenCaso Clinico Oftalmoleyva_777100% (2)
- Pregs NeuroDokument48 SeitenPregs NeuromariancandelariacanizaNoch keine Bewertungen
- Informe 8Dokument5 SeitenInforme 8Carlos Andres GNoch keine Bewertungen
- Taller 3. Cultivo Celular - 2020-Ii PDFDokument2 SeitenTaller 3. Cultivo Celular - 2020-Ii PDFDayana ChicaNoch keine Bewertungen
- Cuestionario de EgoDokument15 SeitenCuestionario de EgoMel alixon Farell AguilarNoch keine Bewertungen
- Ingeniería Genética: antecedentes clave y aplicaciones futurasDokument17 SeitenIngeniería Genética: antecedentes clave y aplicaciones futurasAlejandro TostadoNoch keine Bewertungen
- Topografia Del Maxilar InferiorDokument12 SeitenTopografia Del Maxilar Inferiorcarina nasifNoch keine Bewertungen
- Lectura 2017-6 ZelekDokument15 SeitenLectura 2017-6 ZelekYolanda HernandezNoch keine Bewertungen
- Grupo 5Dokument20 SeitenGrupo 5sijnhNoch keine Bewertungen
- Areas Libres de ConflictoDokument1 SeiteAreas Libres de ConflictoAnonymous fWD7vnGh1Noch keine Bewertungen
- Claves Taxonomicas Serpientes Morfo Fauna 2021Dokument22 SeitenClaves Taxonomicas Serpientes Morfo Fauna 2021LAURA CAROLINA FRANCO MEJIANoch keine Bewertungen