Das könnte Ihnen auch gefallen
- Agua Oxigenada Un Liquido MilagrosoDokument6 SeitenAgua Oxigenada Un Liquido MilagrosoFidel Cienfuegos Valencia100% (1)
- Compuestos NitrogenadosDokument192 SeitenCompuestos NitrogenadosMilton IsraelNoch keine Bewertungen
- 2016-III Simulacro 06.03 CompletoDokument11 Seiten2016-III Simulacro 06.03 CompletoRoger Moreyra QuinchoNoch keine Bewertungen
- Clase 4 FISIOLOGÍA DE LA SANGRE 1Dokument59 SeitenClase 4 FISIOLOGÍA DE LA SANGRE 1Maximo Salazar100% (1)
- Anatomia Fisiologia e Higiene PDFDokument145 SeitenAnatomia Fisiologia e Higiene PDFjorgeluischato100% (1)
- ArteroesclerosisDokument72 SeitenArteroesclerosisKatherine GallosoNoch keine Bewertungen
- Ortodoncia PrequirugicaDokument48 SeitenOrtodoncia PrequirugicaSergio Llufire MendozaNoch keine Bewertungen
- PDF 1 - Biodescodificacion - PaleoliticoDokument67 SeitenPDF 1 - Biodescodificacion - PaleoliticocamelieNoch keine Bewertungen
- Metabolismo CatabolismoDokument33 SeitenMetabolismo CatabolismoFrancis A. Batarse0% (1)
- ENZIMAS bIOQUÍMICADokument37 SeitenENZIMAS bIOQUÍMICANoelis LimonNoch keine Bewertungen
- Comportamiento IndividualDokument36 SeitenComportamiento IndividualBryan Guerra50% (2)
- Sangre y HemostasiaDokument57 SeitenSangre y HemostasiaMacarena Riffo WestermannNoch keine Bewertungen
- Bioquimica Proteinas Plasmaticas TerminadoDokument15 SeitenBioquimica Proteinas Plasmaticas Terminadoerick marioNoch keine Bewertungen
- Guia de Control de Calidad Interno y Externo OkDokument18 SeitenGuia de Control de Calidad Interno y Externo OkJorge GarciaNoch keine Bewertungen
- 1.3 Control Genético de La Síntesis Proteica, Las Funciones deDokument34 Seiten1.3 Control Genético de La Síntesis Proteica, Las Funciones deDF100% (1)
- EosinófilosDokument7 SeitenEosinófilosChristy Sandoval100% (1)
- Sistema NerviosoDokument51 SeitenSistema NerviosoJuan Carlos Serrano TiradoNoch keine Bewertungen
- Rosales FinalDokument28 SeitenRosales FinalLesly ResendizNoch keine Bewertungen
- Metabolismo Lìpidos 21-2Dokument90 SeitenMetabolismo Lìpidos 21-2Joel PMNoch keine Bewertungen
- 13) Reticulo EndoplasmicoDokument27 Seiten13) Reticulo EndoplasmicoPerla Gutiérrez RogelNoch keine Bewertungen
- Clase de Topografía Toracopleural IvDokument34 SeitenClase de Topografía Toracopleural Ivstefanct6421Noch keine Bewertungen
- RibosomasDokument12 SeitenRibosomasCiro PeruNoch keine Bewertungen
- Tejido Sanguineo - GlobulosDokument25 SeitenTejido Sanguineo - GlobulosKevin Flores100% (3)
- Funciones Del AmpcDokument3 SeitenFunciones Del AmpcRaulNoch keine Bewertungen
- MORFOLOGÍA Y ESTRUCTURA BACTERIANA II Estudiantes PDFDokument56 SeitenMORFOLOGÍA Y ESTRUCTURA BACTERIANA II Estudiantes PDFlordgimli77Noch keine Bewertungen
- Primera Clase de BiologiaDokument28 SeitenPrimera Clase de BiologiaLuxa Valverde ChicllaNoch keine Bewertungen
- Carbohidratos 1 2013 Clase Unidad IDokument63 SeitenCarbohidratos 1 2013 Clase Unidad ICecilio Silva LobatonNoch keine Bewertungen
- Metabolismo de Otros Azucares Parte 3Dokument15 SeitenMetabolismo de Otros Azucares Parte 3Daniela Gomez100% (1)
- Transporte de ProteinasDokument9 SeitenTransporte de ProteinasGise MathierNoch keine Bewertungen
- Cuestionario de BioquimicaDokument8 SeitenCuestionario de BioquimicaBel OcNoch keine Bewertungen
- Sindrome Motor EspiramidalDokument21 SeitenSindrome Motor EspiramidalLuis HerreraNoch keine Bewertungen
- 01 - Sangre - Suero y PlasmaDokument20 Seiten01 - Sangre - Suero y PlasmaInés Carmen PastrianNoch keine Bewertungen
- Replicacion Del ADNDokument2 SeitenReplicacion Del ADNCieloDelCarpioNoch keine Bewertungen
- AnemiasDokument79 SeitenAnemiasPinduisaca Luis100% (1)
- Inhibidor EnzimáticoDokument18 SeitenInhibidor EnzimáticoAdri CruzNoch keine Bewertungen
- Procariotas y VirusDokument19 SeitenProcariotas y Viruszonahiphop82% (11)
- Aparato GolgiDokument12 SeitenAparato GolgiSoraida Sanchez Parra0% (1)
- Adaptación Del Sistema HEPATICODokument42 SeitenAdaptación Del Sistema HEPATICOMaría Angelica DAppollonioNoch keine Bewertungen
- Aparato de Golgi - Docx 22Dokument6 SeitenAparato de Golgi - Docx 22Hoa Oscori AlmarazNoch keine Bewertungen
- 1.5 Nucleósidos y NucleótidosDokument40 Seiten1.5 Nucleósidos y NucleótidosKenneth CruzNoch keine Bewertungen
- Transcripcion Del AdnDokument2 SeitenTranscripcion Del Adnflorencia barrientosNoch keine Bewertungen
- Clase 10 CitoesqueletoDokument60 SeitenClase 10 CitoesqueletoElizabethMartinezBeltranNoch keine Bewertungen
- 4.3 GlobosidosDokument10 Seiten4.3 GlobosidosJosé María MurilloNoch keine Bewertungen
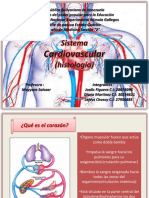
- Sistema Cardiovascular HistologiaDokument26 SeitenSistema Cardiovascular HistologiaAlbany ArteagaNoch keine Bewertungen
- Diapositiva de Reticulo Endoplasmatico LisoDokument8 SeitenDiapositiva de Reticulo Endoplasmatico LisoDaniela Monserrat Olarte SalazarNoch keine Bewertungen
- Adhesion CelularDokument30 SeitenAdhesion CelularBeto HdzNoch keine Bewertungen
- Clase 26 Lipogenesis Ci 2019 PDFDokument42 SeitenClase 26 Lipogenesis Ci 2019 PDFJorge SalomonNoch keine Bewertungen
- Cuestionario de BioquímicaDokument8 SeitenCuestionario de BioquímicaAlejandro NarvaezNoch keine Bewertungen
- Filtrado GlomerularDokument9 SeitenFiltrado GlomerularSabrina SouzaNoch keine Bewertungen
- Código GenéticoDokument18 SeitenCódigo Genéticopaz_saavedra_3Noch keine Bewertungen
- T4 Enfermedades-GlomerularesDokument83 SeitenT4 Enfermedades-GlomerularesKrishna Carmela Janampa ArangoNoch keine Bewertungen
- 5 - SuprarrenalDokument50 Seiten5 - SuprarrenalMirko Miguel Quispe ChumbesNoch keine Bewertungen
- Retículo Endoplásmico Pi MZVDokument29 SeitenRetículo Endoplásmico Pi MZVCool BoyLVNoch keine Bewertungen
- Las Enzimas PDFDokument9 SeitenLas Enzimas PDFTriny VilladiegoNoch keine Bewertungen
- RECEPTORESDokument5 SeitenRECEPTORESMaryen Gallardo ovandoNoch keine Bewertungen
- Taller Metabolismo de Lípidos IIDokument3 SeitenTaller Metabolismo de Lípidos IIAndrés Julián Londoño Cadavid0% (1)
- Bioqumica2 140307082934 Phpapp01Dokument21 SeitenBioqumica2 140307082934 Phpapp01Julimar FloresNoch keine Bewertungen
- Practica Vasos Sanguineos LlenoDokument4 SeitenPractica Vasos Sanguineos LlenoBioqui Mica100% (1)
- HormonasDokument13 SeitenHormonasCarlos Janampa RogelioNoch keine Bewertungen
- ELECTROLITOs Examen ClínicoDokument33 SeitenELECTROLITOs Examen ClínicoSARA VIRIDIANA ALVAREZ MARTINNoch keine Bewertungen
- GluconeogénesisDokument14 SeitenGluconeogénesisChiara María Girardi MarmolNoch keine Bewertungen
- Sintesis de NucleotidosDokument18 SeitenSintesis de NucleotidosErick A. ZepedaNoch keine Bewertungen
- Toxoplasmosis Caso ClinicoooDokument9 SeitenToxoplasmosis Caso ClinicoooJezabel HuamanNoch keine Bewertungen
- Seminario FisioDokument27 SeitenSeminario FisioSteffanie LopezNoch keine Bewertungen
- CLASE 3 Control PostraduccionalDokument47 SeitenCLASE 3 Control PostraduccionalmelissaNoch keine Bewertungen
- NUCLEO CELULAR Cromatina y CromosomasDokument44 SeitenNUCLEO CELULAR Cromatina y CromosomasAbraham CutipaNoch keine Bewertungen
- Bloque Tematico 2 BiologiaDokument39 SeitenBloque Tematico 2 Biologiaangelica carrilloNoch keine Bewertungen
- Dogma Central de La Biología MolecularDokument35 SeitenDogma Central de La Biología Moleculargualterio75Noch keine Bewertungen
- Herpetofaunade MéxicocentralDokument241 SeitenHerpetofaunade MéxicocentralLesly ResendizNoch keine Bewertungen
- Act.1 U1 RSMEDokument7 SeitenAct.1 U1 RSMELesly ResendizNoch keine Bewertungen
- Diversidad de Reptiles en El Cerro Del XitleDokument7 SeitenDiversidad de Reptiles en El Cerro Del XitleLesly ResendizNoch keine Bewertungen
- Duran Et Al 2009 Arañas Urbanas PDFDokument15 SeitenDuran Et Al 2009 Arañas Urbanas PDFLesly ResendizNoch keine Bewertungen
- Centruroides en MéxicoDokument10 SeitenCentruroides en MéxicoLesly ResendizNoch keine Bewertungen
- El Intemperismo en Las Rocas Ígneas y SedimentariasDokument3 SeitenEl Intemperismo en Las Rocas Ígneas y SedimentariasLesly ResendizNoch keine Bewertungen
- Los Mamíferos y Sus 21 Ordenes o GruposDokument10 SeitenLos Mamíferos y Sus 21 Ordenes o GruposLesly Resendiz0% (1)
- ForaminiferosDokument21 SeitenForaminiferosLesly ResendizNoch keine Bewertungen
- Diseccion Del Miembro Superior 2Dokument3 SeitenDiseccion Del Miembro Superior 2Eliana MartinezNoch keine Bewertungen
- Práctica 7 - Determinación de Oxígeno Por El Método de Sensores en Elodea en Condiciones de Luz y OscuridadDokument5 SeitenPráctica 7 - Determinación de Oxígeno Por El Método de Sensores en Elodea en Condiciones de Luz y OscuridadaldairNoch keine Bewertungen
- Reporte de Práctica-FrotisDokument7 SeitenReporte de Práctica-FrotisPamela RomeroNoch keine Bewertungen
- Ajustes de Parámetros en TACDokument4 SeitenAjustes de Parámetros en TACJorge Junior Pereda Ruiz0% (1)
- Caracteristicas GeneralesDokument2 SeitenCaracteristicas GeneralestortillasdemaizNoch keine Bewertungen
- Infecciones Respiratorias AltasDokument59 SeitenInfecciones Respiratorias AltasGonzalo Castillo BarredaNoch keine Bewertungen
- Hiperemesis GravidicaDokument5 SeitenHiperemesis GravidicaValtin KenNoch keine Bewertungen
- Características de Un DiluyenteDokument3 SeitenCaracterísticas de Un DiluyenteluisfranvillalbaNoch keine Bewertungen
- Cuento TroncoDokument2 SeitenCuento TroncoViviana EcheverriNoch keine Bewertungen
- Neoplaisia 2 Cancer de MamaDokument6 SeitenNeoplaisia 2 Cancer de MamaLuis Bojorge100% (1)
- Capítulo 1 Fisiología de Guyton ResumenDokument7 SeitenCapítulo 1 Fisiología de Guyton ResumenValentina BernalNoch keine Bewertungen
- Hernia de Disco Libro Dres GuzmanDokument64 SeitenHernia de Disco Libro Dres GuzmanXochilt BellosoNoch keine Bewertungen
- Informe Eco RIDokument3 SeitenInforme Eco RIMariana OlivaresNoch keine Bewertungen
- Formulas para Calculos Hemodinamicos en PediatriaDokument29 SeitenFormulas para Calculos Hemodinamicos en Pediatriajcguerre100% (3)
- Halógenos y PseudohalógenosDokument13 SeitenHalógenos y PseudohalógenosKarina Cortes0% (1)
- Bco Preguntas Admisión-CTSEDokument8 SeitenBco Preguntas Admisión-CTSELourdes del Socorro Muñoz OrtizNoch keine Bewertungen
- Tronco EncefálicoDokument2 SeitenTronco EncefálicoalextumNoch keine Bewertungen
- Casos Clinicos 3Dokument5 SeitenCasos Clinicos 3capirote73Noch keine Bewertungen
- Artrología CompletoDokument153 SeitenArtrología CompletoLucho GonzalezNoch keine Bewertungen
- Taller Practica IIIDokument11 SeitenTaller Practica IIIMaría CamilaNoch keine Bewertungen
- Fosas Nasales - Lengua y OídoDokument28 SeitenFosas Nasales - Lengua y OídoChristian Ruiz Bustamante100% (1)
- Composicion de La LecheDokument8 SeitenComposicion de La LecheLes SorianoNoch keine Bewertungen
- Artritis Reumatoide y JuvenilDokument29 SeitenArtritis Reumatoide y JuvenilJaviercito BlackNoch keine Bewertungen