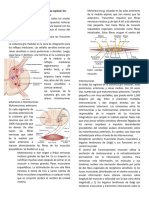
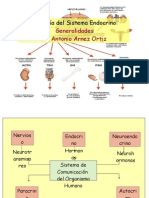
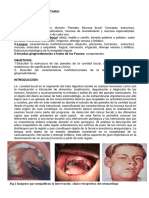
Das könnte Ihnen auch gefallen
- Funciones Motoras de La Médula Espinal2Dokument77 SeitenFunciones Motoras de La Médula Espinal2Erwin RodriguezNoch keine Bewertungen
- Vías AscendentesDokument40 SeitenVías Ascendentescharly_medi100% (2)
- Apunte de Neurofisiologia Del Dr. Eleri Sworres v1Dokument92 SeitenApunte de Neurofisiologia Del Dr. Eleri Sworres v1amadoacosta2013100% (1)
- Sensibilidades Somáticas I. Cap. 47 GuytonDokument24 SeitenSensibilidades Somáticas I. Cap. 47 GuytonVictor Eloy Gonzalez50% (6)
- Capítulo 45 GuytonDokument51 SeitenCapítulo 45 GuytonBillcar Calderon88% (8)
- Medula EspinalDokument64 SeitenMedula EspinalAndres Alba100% (1)
- Bulbo RaquídeoDokument26 SeitenBulbo Raquídeojlcordero100% (1)
- Guyton Vias SensitivasDokument13 SeitenGuyton Vias SensitivassaraNoch keine Bewertungen
- Resumen NeurofisiologíaDokument34 SeitenResumen NeurofisiologíaVirginia MaitreNoch keine Bewertungen
- Corteza Cerebral, Funciones Intelectuales Del CerebroDokument25 SeitenCorteza Cerebral, Funciones Intelectuales Del CerebroMaleni MartínezNoch keine Bewertungen
- Cerebelo y Ganglios BasalesDokument31 SeitenCerebelo y Ganglios BasalesEnrique100% (3)
- Médula EspinalDokument17 SeitenMédula EspinalCARMEN RUBI COLQUE ROBLES100% (1)
- Configuracion Interna Del CerebroDokument6 SeitenConfiguracion Interna Del Cerebroc0raz0nNoch keine Bewertungen
- Medicamentos Inhibidores de La Colinesterasa y Activadores de Los ColinesteroidesDokument5 SeitenMedicamentos Inhibidores de La Colinesterasa y Activadores de Los ColinesteroidesFiodor Tejada ToribioNoch keine Bewertungen
- Cerebelo Microanatomia IDokument29 SeitenCerebelo Microanatomia IAle MedranoNoch keine Bewertungen
- Corteza CerebralDokument25 SeitenCorteza CerebralAlejandra Garcia ObregonNoch keine Bewertungen
- Los Sentidos EspecialesDokument25 SeitenLos Sentidos Especialessulin daniela puerto ovando100% (1)
- Medula EspinalDokument13 SeitenMedula Espinalpollodoro0% (1)
- Configuracion Externa Del CerebroDokument16 SeitenConfiguracion Externa Del CerebroYiro G Nakamura Cardemil100% (1)
- Sensibilidad Somática (Fisiología)Dokument47 SeitenSensibilidad Somática (Fisiología)Jeniffer Scarlett Velasco Bravo100% (6)
- Sustancia Negra, Blanca y GrisDokument7 SeitenSustancia Negra, Blanca y Grisabdiel liraNoch keine Bewertungen
- Cerebelo y Control Motor GeneralDokument40 SeitenCerebelo y Control Motor GeneralEvely OliveiraNoch keine Bewertungen
- Tronco EncefálicoDokument21 SeitenTronco EncefálicoJavier VelasquezNoch keine Bewertungen
- Tálamo e HipotálamoDokument13 SeitenTálamo e HipotálamoVeimar Cordova100% (1)
- Neurofisiologia Del OjoDokument49 SeitenNeurofisiologia Del OjoIdaliiBOo100% (1)
- Ganglios BasalesDokument46 SeitenGanglios BasalesAndres DangondNoch keine Bewertungen
- Anatomía Interna Del Tallo CerebralDokument27 SeitenAnatomía Interna Del Tallo Cerebraljose NavasNoch keine Bewertungen
- Ganglios BasalesDokument35 SeitenGanglios BasalesMFLL - UDCHNoch keine Bewertungen
- Control de Funcion Motora Por Corteza y Tronco Encefalico-1Dokument36 SeitenControl de Funcion Motora Por Corteza y Tronco Encefalico-1sharonpaolaNoch keine Bewertungen
- NeurotransmisoresDokument8 SeitenNeurotransmisores110899Noch keine Bewertungen
- El Sistema Nervioso Autónomo y La Médula SuprarrenalDokument3 SeitenEl Sistema Nervioso Autónomo y La Médula SuprarrenalPablo Soto OsunaNoch keine Bewertungen
- Capítulo 17 Ganglios BasalesDokument13 SeitenCapítulo 17 Ganglios BasalesJochiJaramilloNoch keine Bewertungen
- Codificacion SensorialDokument10 SeitenCodificacion SensorialEiliana Santana100% (1)
- Neuroanatomía - Cap 8. Medula EspinalDokument19 SeitenNeuroanatomía - Cap 8. Medula Espinalwww.pacourgencias.blogspot.com/100% (2)
- El TelencéfaloDokument16 SeitenEl TelencéfaloPerla Lizbeth GuijarroNoch keine Bewertungen
- Ganglios BasalesDokument4 SeitenGanglios BasalesIgnacio Luis Barria Olmedo0% (1)
- Tabla de NeurotransmisoresDokument3 SeitenTabla de NeurotransmisoresAle Ávalos LeónNoch keine Bewertungen
- CEREBELO ResumenDokument8 SeitenCEREBELO ResumenMelina BarviNoch keine Bewertungen
- Caracteres Sexuales Primarios Femeninos y MasculinosDokument5 SeitenCaracteres Sexuales Primarios Femeninos y MasculinosDportivoTlashco50% (4)
- R Talamo SalazarDokument46 SeitenR Talamo SalazarLemaje Leyva RamirezNoch keine Bewertungen
- Neurotrasmisores y NeuropeptidosDokument42 SeitenNeurotrasmisores y NeuropeptidosYudira Chacon PalominoNoch keine Bewertungen
- Sistema Nervioso Autónomo y La Medula Suprarrenal Cap 60Dokument45 SeitenSistema Nervioso Autónomo y La Medula Suprarrenal Cap 60Grhizzel ValladaresNoch keine Bewertungen
- Alteraciones de La ConcienciaDokument75 SeitenAlteraciones de La ConcienciaEvelyn Espinoza AlianoNoch keine Bewertungen
- Medula EspinalDokument44 SeitenMedula EspinalDavidSumerenteTorres100% (1)
- Vias Ascendentes y Descendentes LAMINAS de REDEXDokument22 SeitenVias Ascendentes y Descendentes LAMINAS de REDEXvalentina garcia solanoNoch keine Bewertungen
- Corteza CerebralDokument35 SeitenCorteza CerebralGiovanna Uceda60% (5)
- La Neurona y Sus PartesDokument2 SeitenLa Neurona y Sus PartesNicole Barrera100% (1)
- Clase10-Corteza Cerebral Funciones Intelectuales AprendizajeDokument42 SeitenClase10-Corteza Cerebral Funciones Intelectuales AprendizajeDANIEL ALEJANDRO83% (6)
- SinapsisDokument12 SeitenSinapsisFiorella Davila Sanchez0% (1)
- Diferencia Entre El Sistema Nervioso Simpático y ParasimpáticoDokument5 SeitenDiferencia Entre El Sistema Nervioso Simpático y ParasimpáticodiegoNoch keine Bewertungen
- Anatomia y Fisiologia Del Sistema Nervioso CentralDokument13 SeitenAnatomia y Fisiologia Del Sistema Nervioso Centralmanuel briñezNoch keine Bewertungen
- Primer Par CranealDokument30 SeitenPrimer Par CranealWaleska Aguilar GomezNoch keine Bewertungen
- Ganglios BasalesDokument9 SeitenGanglios BasalesTainy SanchezNoch keine Bewertungen
- Corteza CerebralDokument34 SeitenCorteza CerebralAna LaresNoch keine Bewertungen
- Ganglios BasalesDokument21 SeitenGanglios BasalesDANIEL ALEJANDRONoch keine Bewertungen
- DiencefaloDokument8 SeitenDiencefaloWicho SpacesNoch keine Bewertungen
- Funcion Motora MedularDokument53 SeitenFuncion Motora MedularAlejandro D. Pérez GuillénNoch keine Bewertungen
- Capítulo 55Dokument5 SeitenCapítulo 55Cynthia GonzálezNoch keine Bewertungen
- Resumen Capitulo 55Dokument8 SeitenResumen Capitulo 55Rashel MoscosoNoch keine Bewertungen
- Capítulo 54 - GuytonDokument4 SeitenCapítulo 54 - GuytonJesica AndanNoch keine Bewertungen
- Trauma Ocular y NasalDokument43 SeitenTrauma Ocular y NasaljosemerelloNoch keine Bewertungen
- Seminarios - Fisiologia I 2019Dokument102 SeitenSeminarios - Fisiologia I 2019josemerelloNoch keine Bewertungen
- Funciones Secretoras Del Tubo DigestivoDokument100 SeitenFunciones Secretoras Del Tubo DigestivojosemerelloNoch keine Bewertungen
- GESTION DEL RIESGO EN SALUD Dr. MerelloDokument23 SeitenGESTION DEL RIESGO EN SALUD Dr. Merellojosemerello100% (1)
- HERRAMIENTAS PARA LA CALIDAD Dr. MerelloDokument19 SeitenHERRAMIENTAS PARA LA CALIDAD Dr. MerellojosemerelloNoch keine Bewertungen
- Fisiologia Del GustoDokument22 SeitenFisiologia Del GustojosemerelloNoch keine Bewertungen
- Fisiologia Del Higado y Vesicula Biliar Dr. MerelloDokument32 SeitenFisiologia Del Higado y Vesicula Biliar Dr. MerellojosemerelloNoch keine Bewertungen
- Plan SAR PeruDokument15 SeitenPlan SAR PerujosemerelloNoch keine Bewertungen
- Libro de Bolsillo AHA 2010Dokument107 SeitenLibro de Bolsillo AHA 2010Farid RodríguezNoch keine Bewertungen
- Banco de PreguntasDokument215 SeitenBanco de PreguntasjosemerelloNoch keine Bewertungen
- Sistema EndocrinoDokument81 SeitenSistema EndocrinojosemerelloNoch keine Bewertungen
- Cartilla de Gestion Reactiva Del RiesgoDokument46 SeitenCartilla de Gestion Reactiva Del RiesgojosemerelloNoch keine Bewertungen
- Exposicion de Alteraciones Del PotasioDokument38 SeitenExposicion de Alteraciones Del Potasiojosemerello100% (1)
- Triptico Sistema Respiratorio 130130192924 Phpapp02Dokument2 SeitenTriptico Sistema Respiratorio 130130192924 Phpapp02एलेक्स विगोNoch keine Bewertungen
- Copia de Mapa Conceptual RosaDokument1 SeiteCopia de Mapa Conceptual RosaLUCYEL ALEXANDRA FLORES CHATANoch keine Bewertungen
- Sistema Endocrino - GeneralidadesDokument26 SeitenSistema Endocrino - GeneralidadesJosé Eduardo Pinheiro MatosNoch keine Bewertungen
- Guia 4 Unidad 3 El Corazon y Su Organizacion InternaDokument13 SeitenGuia 4 Unidad 3 El Corazon y Su Organizacion InternaDaniela Valeria MorlaNoch keine Bewertungen
- 2702-LIBRO N°3 - SALUD HUMANA-BIOLOGÍA MENCIÓN-2016-web PDFDokument64 Seiten2702-LIBRO N°3 - SALUD HUMANA-BIOLOGÍA MENCIÓN-2016-web PDFMACARENA CASTILLO MUNOZNoch keine Bewertungen
- Up 1Dokument95 SeitenUp 1juanaNoch keine Bewertungen
- Visceras Pelvicas PDFDokument29 SeitenVisceras Pelvicas PDFJatniel DelgadoNoch keine Bewertungen
- FlagelosDokument10 SeitenFlagelosIrving IvanNoch keine Bewertungen
- Liquido Amniotico PDFDokument4 SeitenLiquido Amniotico PDFMaria Alejandra Guzmán Quijada100% (1)
- 8-Tejido Cartilaginoso.Dokument41 Seiten8-Tejido Cartilaginoso.Zoe RivarolaNoch keine Bewertungen
- Resumen Completo de Histolog¡ADokument105 SeitenResumen Completo de Histolog¡ADaniel RicoNoch keine Bewertungen
- Vias ColinergicasDokument18 SeitenVias Colinergicasfattyev33% (3)
- Alumbramiento Normal y PatológicoDokument6 SeitenAlumbramiento Normal y PatológicoAlexisNoch keine Bewertungen
- BienvenidosDokument26 SeitenBienvenidosElizabeth Garcia FloresNoch keine Bewertungen
- Taller Biología Grado 4 SolucionadoDokument5 SeitenTaller Biología Grado 4 SolucionadoWILLIAMESPINOSANoch keine Bewertungen
- Reforzamiento Aparato Digestivo PDFDokument8 SeitenReforzamiento Aparato Digestivo PDFMarco Antonio Condori DianderasNoch keine Bewertungen
- Resumen Cavidad Oral Apuntes de ClaseDokument8 SeitenResumen Cavidad Oral Apuntes de ClaseBelen SaezNoch keine Bewertungen
- FX Celular FisiopatologiaDokument34 SeitenFX Celular Fisiopatologiamarta parraNoch keine Bewertungen
- Act. 35-36 MC.5. Cavidad BucalDokument23 SeitenAct. 35-36 MC.5. Cavidad BucalEsmil RocaNoch keine Bewertungen
- Vías NasalesDokument1 SeiteVías NasalesChristian Mareco Fernandez100% (1)
- Tejido EpitelialDokument6 SeitenTejido EpitelialNicole BarreraNoch keine Bewertungen
- Desarrollo EmbrionalDokument7 SeitenDesarrollo EmbrionalMariaNoch keine Bewertungen
- Respuestas Cuestionario OrtodonciaDokument48 SeitenRespuestas Cuestionario OrtodonciaPabloAndrésAstudilloNoch keine Bewertungen
- Tema 1 - Anato e Histo IIDokument2 SeitenTema 1 - Anato e Histo IIJuli ColomboNoch keine Bewertungen
- Sistema MuscularDokument14 SeitenSistema MuscularErick Rosales CerpaNoch keine Bewertungen
- Niño Con Parálisis Por Lesión Periférica Al NacimientoDokument70 SeitenNiño Con Parálisis Por Lesión Periférica Al NacimientoLourdes Berenice Garfias SorianoNoch keine Bewertungen
- Tríptico Anatomia PalpatoriaDokument2 SeitenTríptico Anatomia PalpatoriaCarol Mata BarreraNoch keine Bewertungen
- Control Cardiovascular 2015Dokument29 SeitenControl Cardiovascular 2015CarlosIvánSánchezGuerreroNoch keine Bewertungen
- Programa Anatomía ESM IPNDokument46 SeitenPrograma Anatomía ESM IPNJesus Ubaldo Peñaloza100% (1)
- Tesis Final 5.0 Original + Indice GeneralDokument100 SeitenTesis Final 5.0 Original + Indice GeneralJonathan Elias Barrientos OssesNoch keine Bewertungen