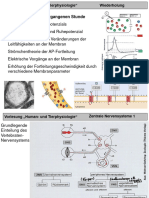
Das könnte Ihnen auch gefallen
- Nelles Guide Reiseführer Baltische Staaten: Estland, Lettland, Litauen, KaliningradVon EverandNelles Guide Reiseführer Baltische Staaten: Estland, Lettland, Litauen, KaliningradNoch keine Bewertungen
- Physiologie 3 230424 164353Dokument39 SeitenPhysiologie 3 230424 164353AcuNoch keine Bewertungen
- M Q2 LK Tafelbilder 15-3-24Dokument4 SeitenM Q2 LK Tafelbilder 15-3-24saleahshah2006Noch keine Bewertungen
- Gleichschenkliges: RechtwinkligesDokument1 SeiteGleichschenkliges: RechtwinkligesHiloloNoch keine Bewertungen
- Analytische Geometrie 2Dokument5 SeitenAnalytische Geometrie 2ManukaaaNoch keine Bewertungen
- MessTechnik VorlesungDokument37 SeitenMessTechnik Vorlesungsoswpg1Noch keine Bewertungen
- 4.4 PotenzreihenansatzDokument2 Seiten4.4 Potenzreihenansatzgimmegimme645Noch keine Bewertungen
- Anonymus - La Corralera (Milonga Campera)Dokument2 SeitenAnonymus - La Corralera (Milonga Campera)Jeanpierre CarabasNoch keine Bewertungen
- Prozess 2017-1Dokument8 SeitenProzess 2017-1wipelop451Noch keine Bewertungen
- AB Säuren Und Säurenrest - IonenDokument1 SeiteAB Säuren Und Säurenrest - Ionenmimirose564Noch keine Bewertungen
- Mozart Guglielma PDFDokument6 SeitenMozart Guglielma PDFAnna WilkNoch keine Bewertungen
- AnleitungDokument64 SeitenAnleitungAz SymNoch keine Bewertungen
- Calciumnitrat Calciumsulfat Eisenchloridlösung Kaliumhydrogenphosphat Kaliumnitrat MagnesiumsulfatDokument1 SeiteCalciumnitrat Calciumsulfat Eisenchloridlösung Kaliumhydrogenphosphat Kaliumnitrat MagnesiumsulfattomNoch keine Bewertungen
- Grund: Eager DerDokument34 SeitenGrund: Eager DerSİNEM ERAYVAZNoch keine Bewertungen
- Pathfinder CharakterbogenDokument1 SeitePathfinder CharakterbogenGustavNoch keine Bewertungen
- ARSA - S.25 - Römische Geschichte Von Erbauung Der - STDokument541 SeitenARSA - S.25 - Römische Geschichte Von Erbauung Der - STPpš MiroNoch keine Bewertungen
- Infoflyer Linie 50Dokument2 SeitenInfoflyer Linie 50423egeNoch keine Bewertungen
- Fahrpläne Stadtwerke AugsburgDokument2 SeitenFahrpläne Stadtwerke AugsburgValentino StellaNoch keine Bewertungen
- Uebungsblatt IT Grundlagen InformatikDokument2 SeitenUebungsblatt IT Grundlagen InformatikluNoch keine Bewertungen
- WarMachine 100Dokument22 SeitenWarMachine 100ady50% (2)
- Diagrama Dimensionare Pardoseala Radianta Uponor Tecto - Comfort-Pipe Plus 17x2Dokument1 SeiteDiagrama Dimensionare Pardoseala Radianta Uponor Tecto - Comfort-Pipe Plus 17x2Stelian BerceaNoch keine Bewertungen
- Trumpet Scales, HighDokument1 SeiteTrumpet Scales, HighScott JohnsonNoch keine Bewertungen
- ULS12000Dokument1 SeiteULS12000Victor AcunaNoch keine Bewertungen
- Briefkopie Mercedes-Benz O 405 G - 068Dokument1 SeiteBriefkopie Mercedes-Benz O 405 G - 068Autogara BacauNoch keine Bewertungen
- Mosque CoordinatesDokument1 SeiteMosque CoordinateseiconszayedNoch keine Bewertungen
- DeutschDokument152 SeitenDeutschfree vpsNoch keine Bewertungen
- Arnold 2350 - 2351 - 2353 - 2354 - FF - E (DB BR 103 Neuer Motor)Dokument2 SeitenArnold 2350 - 2351 - 2353 - 2354 - FF - E (DB BR 103 Neuer Motor)harryblnNoch keine Bewertungen
- Mathe LernzettelDokument11 SeitenMathe Lernzettelfriederike.srlNoch keine Bewertungen
- (2021-04-11T22 - 15 - 17) 4.11. Wegbeschreibung - OptimizedDokument4 Seiten(2021-04-11T22 - 15 - 17) 4.11. Wegbeschreibung - OptimizediremNoch keine Bewertungen
- WSW Fahrplan S Bahn Linie S8Dokument17 SeitenWSW Fahrplan S Bahn Linie S8MikhayiNoch keine Bewertungen
- S8Dokument17 SeitenS8MarkoCimbaljevicNoch keine Bewertungen
- Merkblatt QFDokument1 SeiteMerkblatt QFRenata BotosNoch keine Bewertungen
- T1940 Fortius Trainer 06Dokument2 SeitenT1940 Fortius Trainer 06Rodolfo PimentelNoch keine Bewertungen
- Big Band TuneDokument2 SeitenBig Band TuneJuanvi Lladosa ArnauNoch keine Bewertungen
- Tapcon 230 Expert 6655744 enDokument2 SeitenTapcon 230 Expert 6655744 enLuis AravenaNoch keine Bewertungen
- Kabine + 1010300005962 - 1Dokument1 SeiteKabine + 1010300005962 - 1Mihai CroitoruNoch keine Bewertungen
- DocumentDokument3 SeitenDocument41834Noch keine Bewertungen
- GA 1 Crane Weight and DimensionsDokument3 SeitenGA 1 Crane Weight and DimensionsminhtranNoch keine Bewertungen
- Wagner - Liebestod PG314 SOURCEDokument8 SeitenWagner - Liebestod PG314 SOURCESasko TemelkoskiNoch keine Bewertungen
- Ept 403-53XXX-5X C 05Dokument3 SeitenEpt 403-53XXX-5X C 05Michael MitchellNoch keine Bewertungen
- HandoutDokument9 SeitenHandoutMaxima GetahunNoch keine Bewertungen
- Ept 404-52XXX-5X C 03Dokument2 SeitenEpt 404-52XXX-5X C 03Muhammad RafiqueNoch keine Bewertungen
- 4.3 Lineare DGLN Höherer OrdnungDokument5 Seiten4.3 Lineare DGLN Höherer Ordnunggimmegimme645Noch keine Bewertungen
- To 7Dokument1 SeiteTo 7Trần NghiêmNoch keine Bewertungen
- BA 2180762-200-Zusammenbau TFW 300 Mit 2 PB 200 Schnecken - IdwDokument1 SeiteBA 2180762-200-Zusammenbau TFW 300 Mit 2 PB 200 Schnecken - IdwMatias AcunaNoch keine Bewertungen
- Act 1Dokument1 SeiteAct 1luar 11Noch keine Bewertungen
- Mediamarkt Apple Watch Se Mitt CellDokument1 SeiteMediamarkt Apple Watch Se Mitt Cellchuj chujNoch keine Bewertungen
- FWS 50Dokument1 SeiteFWS 50Artm YakovlevNoch keine Bewertungen
- Ein Koffer SprichtDokument3 SeitenEin Koffer Spricht8b7w9jkdftNoch keine Bewertungen
- Singer 215 (Deutch) Sewing Machine Instruction ManualDokument21 SeitenSinger 215 (Deutch) Sewing Machine Instruction ManualiliiexpugnansNoch keine Bewertungen
- 26.D - Matarqocha UpDokument1 Seite26.D - Matarqocha UpFRANCO VITO MORALES MENACHONoch keine Bewertungen
- Rückholrolle Return Roll: HS Assembly Drawing Baugruppenzeichnung HSDokument1 SeiteRückholrolle Return Roll: HS Assembly Drawing Baugruppenzeichnung HSBui LE VANNoch keine Bewertungen
- BOSCH Connect-Key K 30 RF - Regel - 7736603499 - InstallationDokument84 SeitenBOSCH Connect-Key K 30 RF - Regel - 7736603499 - InstallationSkala55Noch keine Bewertungen
- Notizen VWL 11111Dokument1 SeiteNotizen VWL 11111Bedo khaledNoch keine Bewertungen
- Wiring Diagram Central Door Lock Mobil Avanzaxenia Tahun 2006Dokument2 SeitenWiring Diagram Central Door Lock Mobil Avanzaxenia Tahun 2006Mardiansyah Black100% (3)
- Aus Dem Viljaet KosovoDokument1 SeiteAus Dem Viljaet KosovopoposkidimitarNoch keine Bewertungen
- Confirmation PDFDokument2 SeitenConfirmation PDFAdela MarasescuNoch keine Bewertungen
- Tygel, David - Café Com LeiteDokument3 SeitenTygel, David - Café Com LeitePierre F.Noch keine Bewertungen
- AnalysisDokument11 SeitenAnalysisRiad K.Noch keine Bewertungen
- Vivaldi - FrühlingDokument8 SeitenVivaldi - Frühlinganže RupnikNoch keine Bewertungen
- Physiologie 5 230504 134705Dokument28 SeitenPhysiologie 5 230504 134705AcuNoch keine Bewertungen
- Physiologie 6 230419 114748Dokument28 SeitenPhysiologie 6 230419 114748AcuNoch keine Bewertungen
- VL13 Zellorganellen 2 Mitochondrien Chloroplasten SkriptDokument31 SeitenVL13 Zellorganellen 2 Mitochondrien Chloroplasten SkriptAcuNoch keine Bewertungen
- VL12 Zellorganellen 1 Kern ER Golgi Proteinsortierung SkriptDokument35 SeitenVL12 Zellorganellen 1 Kern ER Golgi Proteinsortierung SkriptAcuNoch keine Bewertungen
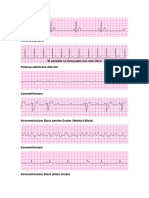
- EKG Interpretation J BereutherDokument9 SeitenEKG Interpretation J BereutherAntonelaDulcik100% (1)
- Sportler EkgDokument2 SeitenSportler EkgNils VerborgNoch keine Bewertungen
- EKGmory HandbuchDokument16 SeitenEKGmory HandbuchAzra ZolicNoch keine Bewertungen
- Acls EkgDokument4 SeitenAcls EkgScribdTranslationsNoch keine Bewertungen
- Analyse Eines EKGDokument6 SeitenAnalyse Eines EKGDaived TrombettaNoch keine Bewertungen
- EKG Grundlagen SkriptDokument22 SeitenEKG Grundlagen SkriptmbarevoNoch keine Bewertungen
- Belastung BeanspruchungDokument21 SeitenBelastung BeanspruchungSebastián HksNoch keine Bewertungen
- EKG-Interpretation: Elektrokardiogramm: Dieser Kurs Vermittelt Ihnen Grundlegende Kenntnisse in Der EKG-InterpretationDokument29 SeitenEKG-Interpretation: Elektrokardiogramm: Dieser Kurs Vermittelt Ihnen Grundlegende Kenntnisse in Der EKG-InterpretationF JNoch keine Bewertungen
- Bedienerhandbuch Zoll M SerieDokument98 SeitenBedienerhandbuch Zoll M SeriePetar RajicNoch keine Bewertungen
- SkyTest® Piloten-Assessment 2024: Handbuch zu den Einstellungstests für Ab-Initio- und Ready-Entry-PilotenVon EverandSkyTest® Piloten-Assessment 2024: Handbuch zu den Einstellungstests für Ab-Initio- und Ready-Entry-PilotenNoch keine Bewertungen
- Sprachbausteine Deutsch B2-C1 Medizin Fachsprachprüfung (FSP): 10 Übungen zur FSP-Prüfungsvorbereitung mit LösungenVon EverandSprachbausteine Deutsch B2-C1 Medizin Fachsprachprüfung (FSP): 10 Übungen zur FSP-Prüfungsvorbereitung mit LösungenBewertung: 3.5 von 5 Sternen3.5/5 (6)
- Pflegekniffe von A - Z: Pflegefehler erfolgreich vermeidenVon EverandPflegekniffe von A - Z: Pflegefehler erfolgreich vermeidenBewertung: 5 von 5 Sternen5/5 (1)
- Handlungsorientierte Pflegedokumentation: Wissen, worauf es ankommtVon EverandHandlungsorientierte Pflegedokumentation: Wissen, worauf es ankommtBewertung: 5 von 5 Sternen5/5 (1)
- Formulierungshilfen für die Strukturierte Informationssammlung SIS und Maßnahmenplanung: Themenfeld Mobilität und BeweglichkeitVon EverandFormulierungshilfen für die Strukturierte Informationssammlung SIS und Maßnahmenplanung: Themenfeld Mobilität und BeweglichkeitNoch keine Bewertungen
- Die Geheimnisse der Visualisierung nutzen lernenVon EverandDie Geheimnisse der Visualisierung nutzen lernenBewertung: 5 von 5 Sternen5/5 (1)
- Die flache Erde oder Hundert Beweise dafür, daß die Erde keine Kugel istVon EverandDie flache Erde oder Hundert Beweise dafür, daß die Erde keine Kugel istNoch keine Bewertungen
- Das bewusste Universum: Wie das Bewusstein die materielle Welt erschafftVon EverandDas bewusste Universum: Wie das Bewusstein die materielle Welt erschafftBewertung: 5 von 5 Sternen5/5 (1)
- Schnelles Denken, langsames Denken. Zusammenfassung & Analyse des Bestsellers von Daniel: Wie Denkfehler die Entscheidungsfindung beeinflussenVon EverandSchnelles Denken, langsames Denken. Zusammenfassung & Analyse des Bestsellers von Daniel: Wie Denkfehler die Entscheidungsfindung beeinflussenBewertung: 5 von 5 Sternen5/5 (2)
- Command Line Kung Fu: Bash-Scripting-Tricks, Linux Tipps und praktische Einzeiler für die ShellVon EverandCommand Line Kung Fu: Bash-Scripting-Tricks, Linux Tipps und praktische Einzeiler für die ShellNoch keine Bewertungen
- Das Sensor-Buch: Mit Elektronik, Arduino und Raspberry Pi die Welt erfassenVon EverandDas Sensor-Buch: Mit Elektronik, Arduino und Raspberry Pi die Welt erfassenNoch keine Bewertungen
- Das neue Werkbuch Elektronik: Das komplette Know-how der Elektronik aktuell erklärtVon EverandDas neue Werkbuch Elektronik: Das komplette Know-how der Elektronik aktuell erklärtBewertung: 5 von 5 Sternen5/5 (1)
- Panzerketten: Die Gleisketten der deutschen Kettenfahrzeuge des Zweiten WeltkriegesVon EverandPanzerketten: Die Gleisketten der deutschen Kettenfahrzeuge des Zweiten WeltkriegesBewertung: 4.5 von 5 Sternen4.5/5 (2)
- Einführung in die Aurachirurgie: Medizin im 21. JahrhundertVon EverandEinführung in die Aurachirurgie: Medizin im 21. JahrhundertNoch keine Bewertungen
- Der Cannabis-Anbau: Alles über Botanik, Anbau, Vermehrung, Weiterverarbeitung und medizinische Anwendung sowie THC-MessverfahrenVon EverandDer Cannabis-Anbau: Alles über Botanik, Anbau, Vermehrung, Weiterverarbeitung und medizinische Anwendung sowie THC-MessverfahrenBewertung: 5 von 5 Sternen5/5 (2)
- DIE FALLE. Was sie ist, wie sie funktioniert und wie wir ihren Illusionen entkommen: »Der Schleier der Illusion wird beiseite gefegt!«Von EverandDIE FALLE. Was sie ist, wie sie funktioniert und wie wir ihren Illusionen entkommen: »Der Schleier der Illusion wird beiseite gefegt!«Noch keine Bewertungen
- Enzyklopädie der Cannabiszucht: Fachbuch der HanfgenetikVon EverandEnzyklopädie der Cannabiszucht: Fachbuch der HanfgenetikBewertung: 5 von 5 Sternen5/5 (1)
- Formulierungshilfen zur Pflegeplanung: Dokumentation der Pflege und Betreuung gemäß Pflegeprozess nach ATL, ABEDL, SIS, Expertenstandards, QPR-Indikatoren und BI des MDKVon EverandFormulierungshilfen zur Pflegeplanung: Dokumentation der Pflege und Betreuung gemäß Pflegeprozess nach ATL, ABEDL, SIS, Expertenstandards, QPR-Indikatoren und BI des MDKNoch keine Bewertungen
- Lehrbuch der Aurachirurgie: Medizin im 21. JahrhundertVon EverandLehrbuch der Aurachirurgie: Medizin im 21. JahrhundertNoch keine Bewertungen
- Sofortheilung durch Borax: Erprobte Anwendungen und dringend notwendiges Praxiswissen für den sicheren Umgang mit Borax PulverVon EverandSofortheilung durch Borax: Erprobte Anwendungen und dringend notwendiges Praxiswissen für den sicheren Umgang mit Borax PulverBewertung: 5 von 5 Sternen5/5 (1)
- Das Geheime Weltraumprogramm der U.S. Navy & Die Allianz mit den Nordischen. Kampftruppen für den Weltraum - die wahren Pläne der Regierung TrumpVon EverandDas Geheime Weltraumprogramm der U.S. Navy & Die Allianz mit den Nordischen. Kampftruppen für den Weltraum - die wahren Pläne der Regierung TrumpNoch keine Bewertungen
- Elektro-Installationen im Haus: Leicht verständliche Anleitungen für völlig unerfahrene Einsteiger und Profi-HeimwerkerVon EverandElektro-Installationen im Haus: Leicht verständliche Anleitungen für völlig unerfahrene Einsteiger und Profi-HeimwerkerNoch keine Bewertungen
- DMT Handbuch - Alles über Dimethyltryptamin, DMT-Herstellungsanleitung und Schamanische PraxistippsVon EverandDMT Handbuch - Alles über Dimethyltryptamin, DMT-Herstellungsanleitung und Schamanische PraxistippsNoch keine Bewertungen
- Tesla: Freie Energie selber bauen Ausgabe 2018 Weiteres Bonusmaterial zum Buch auch auf: https://www.dein-teslabauplan.deVon EverandTesla: Freie Energie selber bauen Ausgabe 2018 Weiteres Bonusmaterial zum Buch auch auf: https://www.dein-teslabauplan.deNoch keine Bewertungen